Клеточное ядро, Хроматин, Ядрышко, Мембрана ядра,
Ядро
text_fields
text_fields
arrow_upward
Ядро — центральный элемент клетки. Его оперативное удаление дискоординирует функции цитоплазмы. Ядро играет главную роль в передаче наследственных признаков и синтезе белков. Передача генетической информации от одной клетки к другой обеспечивается дезоксирибонуклеиновой кислотой (ДНК), содержащейся в хромосомах. Удвоение ДНК предшествует клеточному делению. Масса ядра в клетках разных тканей различна и составляет, например, 10-18% от массы гепатоцита, 60% — в лимфоидных клетках. В интерфазе (межмитотическом периоде) ядро представлено четырьмя элементами: хроматином, нуклеолой (ядрышком), нуклеоплазмой и ядерной мембраной.
Хроматин
text_fields
text_fields
arrow_upward
Для зрелых клеток (например, крови) характерны ядра богатые плотным, конденсированным хроматином, лежащим глыбками. В ядрах соматических клеток женщин он представлен глыбкой хроматина, сближенного с мембраной ядра: это женский половой хроматин (или тельца Барра), представляющий собой конденсированную Х-хромосому. Мужской половой хроматин представлен в ядрах мужских соматических клеток глыбкой, светящейся при окраске флюорохромами. Определение полового хроматина используется, например, для установления пола ребенка по клеткам, полученным из околоплодной жидкости беременной женщины.
Ядрышко
text_fields
text_fields
arrow_upward
Ядрышко — внутриядерная структура сферической формы, не имеющая мембраны. Оно развито во всех клетках, отличающихся высокой активностью белкового синтеза, что связано с образованием в нем субъединиц цитоплазмы, рРНК. Например, ядрышки обнаруживаются в ядрах способных к делению клеток — лимфобластах, миелобластах и др.
Мембрана ядра
text_fields
text_fields
arrow_upward
Мембрана ядра представлена двумя листами, просвет между которыми соединен с полостью эндоплазматического ретикулума. Мембрана имеет отверстия (ядерные поры) приблизительно до 100 нм в диаметре, через которые свободно проходят макромолекулы (рибо-нуклеазы, РНК). Вместе с тем, ядерная мембрана и поры поддерживают микросреду ядра, обеспечивая избирательный обмен различных веществ между ядром и цитоплазмой. В малодифференцированной клетке поры занимают до 10% поверхности ядра, но с созреванием клетки их суммарная поверхность уменьшается.
Нуклеоплазма (ядерный сок)
text_fields
text_fields
arrow_upward
Нуклеоплазма (ядерный сок) представляет собой коллоидный раствор, содержащий белки, который обеспечивает обмен метаболитов и быстрое перемещение молекул РНК к ядерным порам. Количество нуклеоплазмы уменьшается при созревании или старении клетки.
Деление клеток. Митоз.
text_fields
text_fields
arrow_upward
Митоз (рис. 1.5) занимает лишь часть клеточного цикла. В клетках млекопитающих фаза митоза (М) длится около часа.
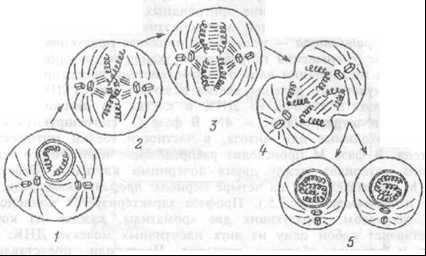 Рис. 1.5 Различные фазы митоза соматической клетки.
Рис. 1.5 Различные фазы митоза соматической клетки.За нею следует постмитотическая пауза (G1), для которой характерна высокая активность биосинтеза белков в клетке, реализуются процессы транскрипции и трансляции. Продолжительность паузы около 10 часов, но это время значительно варьирует и зависит от влияния регулирующих факторов, стимулирующих и тормозящих деление клеток, от снабжения их питательными веществами.
Следующая фаза клеточного цикла характеризуется синтезом (репликацией) ДНК (фаза S) и занимает около 9 часов. Далее следует премитотическая фаза G2, продолжающаяся около 4 часов. Таким образом, весь клеточный цикл длится около 24 часов:
Клетки могут находиться также в фазе покоя — Go, длительно оставаясь вне клеточного цикла. Например, у человека до 90% стволовых кроветворных клеток находится в фазе Go, но их переход из Go в G1 ускоряется при возрастании потребностей в клетках крови.
Высокая чувствительность клеток к регулирующим их деление факторам в фазе G1 объясняется синтезом на мембранах клеток в этот период рецепторов гормонов, стимулирующих и ингибирующих факторов. Например, деление эритроидных клеток костного мозга в фазе G, стимулирует гормон эритропоэтин. Тормозит этот процесс ингибитор эритропоэза — вещество снижающее продукцию эритроцитов в случае уменьшения потребностей тканей в кислороде (глава 6).
Передача информации ядру о взаимодействии рецепторов мембраны со стимулятором деления клетки включает синтез ДНК, т.е. фазу S. В результате количество ДНК в клетке из диплоидного, 2N, переходит в тетраплоидное — 4N. В фазе G2 синтезируются структуры, необходимые для митоза, в частности, белки митотического веретена.
В фазе М происходит распределение идентичного генетического материала между двумя дочерними клетками. Собственно фаза М подразделяется на четыре периода: профазу, метафазу, анафазу и телофазу (рис.1.5.).
Профаза характеризуется конденсацией ДНК хромосом, образующих две хроматиды, каждая из которых представляет собой одну из двух идентичных молекул ДНК. Нуклеола и ядерная оболочка исчезают. Центриоли, представленные тонкими микротрубочками расходятся к двум полюсам клетки, образуя митотическое веретено.
В метафазу хромосомы располагаются в центре клетки, образуя метафазную пластинку, В эту фазу морфология каждой хромосомы наиболее отчетлива, что используется на практике для исследования хромосомного набора клетки.
Анафаза характеризуется движением хроматид, «растаскиваемых» волокнами митотического веретена к противоположным полюсам клетки.
Телофаза характеризуется образованием ядерной мембраны вокруг дочернего набора хромосом. Знание особенностей клеточного цикла используется на практике, например, при создании цитостатических веществ для лечения лейкозов. Так, свойство винкристина быть ядом для митотического веретена используется для остановки митоза лейкозных клеток.
Дифференциация клеток
text_fields
text_fields
arrow_upward
Дифференциация клеток — есть приобретение клеткой специализированных функций, связанное с появлением в ней структур, обес-печивающих выполнение этих функций (например, синтез и накопление гемоглобина в эритроцитах характеризует их дифференциацию в эритроциты). Дифференциация связана с генетически запрограм-мированным торможением (репрессией) функций одних участков генома и активацией других.
Читайте также:
doctor-v.ru
Глава 10. Ядерная оболочка
Структура, ограничивающая параметр клеточного ядра, ядерная оболочка, характерна для эукариотических клеток. Она разделяет два внутриклеточных компартмента друг от друга, цитоплазму от ядра. Значение такого разделения структур в пространстве очень важно: это приводит к обособлению процессов синтеза белка и процессов синтеза нуклеиновых кислот, что создает дополнительные, по сравнению с прокариотами, возможности для регуляции генной активности и ее реализации в виде синтеза специфических белков. Активная регуляция транспорта из цитоплазмы в ядро и из ядра в цитоплазму, через специальные комплексы пор создает систему избирательного транспорта веществ, делая ядерную оболочку “генными воротами” со специальными “превратниками” (контрольными пунктами), регулирующими потоки ядерного импорта и экспорта. Кроме того, как уже описывалось, ядерная оболочка играет большую роль в организации трехмерной структуры интерфазного ядра, элементы ядерной оболочки являются частью ядерного белкового матрикса.Ядерная оболочка состоит из двух мембран, внешней и внутренней, между которыми располагается перинуклеарное пространство (рис. 106). Внутренняя мембрана ядерной оболочки структурно связана с ламиной — фиброзным периферическим слоем ядерного белкового матрикса. В общем виде ядерная оболочка может быть представлена как двухслойный мешок, отделяющий содержимое ядра от цитоплазмы. Однако ядерная оболочка имеет характерную особенность, отличающую ее от других двухмембранных структур клетки (митохондрии и пластиды). Это наличие особых ядерных пор, которые образуются за счет многочисленных зон слияния двух ядерных мембран и представляют собой как бы округлые, сквозные перфорации всей ядерной оболочки.
Компоненты ядерной оболочки
Внешняя мембрана ядерной оболочки
, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматического ретикулума (ЭПР). Так, на внешней ядерной мембране обычно располагается большое количество рибосом, как и на мембранах эргастоплазмы. Существуют многочисленные наблюдения о непосредственном переходе внешней ядерной мембраны в систему каналов эндоплазматического ретикулума, что особенно подчеркивает структурную идентичность этих мембран (рис. 106).Так у клеток, бедных эндоплазматическим ретикулумом, внешняя ядерная мембрана может представлять собой “минимальный” объем эндоплазматического ретикулума, который может участвовать в синтезе белкового и липидного компонентов мембран. Описаны случаи, когда от внешней ядерной мембраны отщепляются мембранные вакуоли, направляющиеся в проксимальный отдел аппарата Гольджи. Состав липидов и белков внешней ядерной мембраны очень схож с таковым ретикулума, что, возможно, и определяет их общие биохимические функции, что особенно подчеркивается наличием рибосом на поверхности мембран, обращенной в гиалоплазму. Эти рибосомы синтезируют, как мембранные, так и секретируемые белки, которые могут транспортироваться в перинуклеарное пространство, а оттуда в полости цистерн ЭПР. Так, например, при стимуляции образования -глобулинов в плазмоцитах первые продукты клеточной активности локализуются в перинуклеарном пространстве, а потом начинают появляться в полостях ЭПР. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность — она может образовывать различной величины выпячивания или выросты в сторону цитоплазмы.
Внутренняя мембрана ядерной оболочки рибосом на своей поверхности не имеет, но связана с фиброзным слоем, ядерной ламиной (Lamina nucleum limitans), которая, в свою очередь, заякоревает хроматин на ядерной оболочке. Связь хроматина с внутренней мембраной оболочки является ее характерной особенностью, хотя существуют примеры, когда эти связи нарушаются при сохранении целостности ядерной оболочки. Так, например, в ооцитах амфибий на стадии диплотены все хромосомы собираются в центре ядра и полностью теряют связь с ядерной оболочкой. С другой стороны, при делении клеток с т.н. закрытым типом митоза большая часть внутренней ядерной мембраны теряет связь с хроматином.
О специфичности белков ламины уже говорилось в разделе “Ядерный белковый матрикс”, здесь же необходимо еще раз подчеркнуть, что эти фибриллярные белки не образуют неизменную структуру. Фиброзный слой ламины все время перестраивается, особенно в связи с ростом поверхности ядра, во время клеточного цикла. Характерные для внутренней ядерной мембраны белки ламины A, C и B относятся к фибриллярным белкам V типа промежуточных филаментов (см. ниже), их фибриллярные мономеры могут образовывать димеры, тетрамеры, а последние образуют фибриллы толщиной около 10 нм. Со стороны кариоплазмы под внутренней ядерной мембраной такие фибриллы образуют ортогональные структуры, чередующиеся с рыхло расположенной сетью этих же фибрилл.
Белки ламины с мембраной связаны двояким образом. Так ламин B после синтеза модифицируется добавлением гидрофобной изопентильной группы вблизи C-конца. Эта липофильная группа встраивается в слой мембраны и как бы заякоревает ламину на мембране. Кроме того целый ряд интегральных белков внутренней ядерной мембраны (LBR, LAR, эмерин и др.) также закрепляют ламины посредством дополнительных белков, входящих в состав этого фиброзного слоя. Эти же белки участвуют в связывании ядерной мембраны с хроматином.
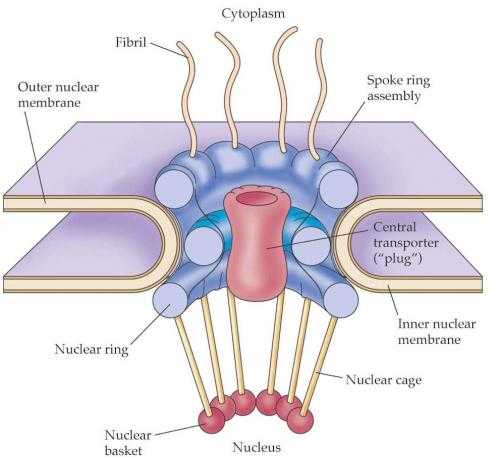
Наиболее характерной и бросающейся в глаза структурой в составе ядерной оболочки является ядерная пора. Поры в оболочке образуются за счет двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром около 100 нм. При альдегидной фиксации или при использовании метода замораживания и скалывания в электронном микроскопе видно, что округлое сквозное отверстие в ядерной оболочке заполнено сложно организованными глобулярными и фибриллярными структурами (рис. 107). Совокупность мембранных перфораций и этих структур называют комплексом пор ядра. Тем самым подчеркивается, что ядерная пора не просто сквозная дыра в ядерной оболочке, через которую непосредственно вещества ядра и цитоплазмы могут сообщаться. Компоненты комплекса пор имеют белковую природу.
Ядерный поровый комплекс (ЯПК или NPC — nuclear pore complex) представляет собой супрамолекулярную структуру с м.в. более 125 х 106 Да, состоящую из более 1000 белков, масса которых в 30 раз больше чем рибосома. Белки ЯПК носят название нуклеопоринов 50-100 видов. Эти белки собраны примерно в 12 субкомплексов.
В последнее время удалось получить отчетливые изображения ЯПК в электронном микроскопе, что дает возможность понять их структурную организацию. Внешний диаметр порового комплекса составляет около 100 нм, а высота — 75 нм. В целом он представляет собой цилиндрическую фигуру с признаками октогональной симметрии. Несмотря на очень впечатляющие изображения выделенных ЯПК, разные авторы дают разные схемы строения этого сложного комплекса, обладающего симметрией восьмого порядка.
Если посмотреть на ЯПК в плане на ультратонком срезе, то бросается в глаза, что его периферия представлена восьмью глобулами (рис. 108, 109). На выделенных же ЯПК в первую очередь видны кольчатые структуры. От периферических компонентов ЯПК в сторону цитоплазмы простираются фибриллярные выросты. Со стороны ядра тоже фибриллярные выросты образуют корзинкоподобную структуру, связанную терминальным кольцом. В большинстве моделей центр цилиндрической фигуры ЯПК содержит “пробку” (центральную гранулу, или транспортер). По одной из моделей (см. рис. 93) цитоплазматические филаменты отходят от цитоплазматического кольца, состоящего из 8 субъединиц. Между ним и внешней ядерной мембраной располагается тонкое кольцо, а затем звездчатое кольцо. Цитоплазматическое кольцо связано внутренними филаментами с транспортером, который находится в центре и заполняет пространство между внешней и внутренней ядерной мембраной. Сходная структура находится на внутренней мембране: нуклеоплазматическое кольцо поддерживает филаменты “корзины”. Другие варианты моделей показаны на (рис. 110).
Весь ЯПК закрепляется интегральными белками, гликопротеидами gp 210 и РОМ 121 в стенке мембранной перфорации.
По своей сложности организации и, главное, по функциональной значимости комплекс ядерной поры можно было бы отнести к органеллам клетки, т.к. их роль заключается в контроле за ядерно-цитоплазменными связями.
Размер ядерных пор и их структура стандартны не только для данной клетки, но и для всех клеток данного организма, более того для всех эукариот.
Число ядерных пор (см. табл. 13) зависит от метаболической активности клеток: чем выше синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра. Так, у эритробластов (клетки-предшественники ядерных эритроцитов) низших позвоночных животных во время интенсивного синтеза и накопления гемоглобина обнаруживается в ядре около 30 ядерных пор на 1 мкм2. После того как эти процессы заканчиваются, в ядрах зрелых клеток — эритроцитов прекращаются синтезы ДНК и РНК, и количество пор падает до 5 на мкм2. В ядерных оболочках полностью зрелых сперматозоидов поры не обнаруживаются, так же как у микронуклеусов некоторых инфузорий. Количество пор может изменяться в течение клеточного цикла. Первое возрастание числа пор наблюдается при реконструкции и росте ядер после митоза, второй этап увеличения числа пор происходит во время синтеза ДНК.
Таблица 13. Количество ядерных пор в различных объектах
Объект | Число ядерных пор на мкм2. | Число пор на одно ядро |
Ксенопус, почки | 10,05 | 3400 |
Ксенопус, ооцит | 51,0 | 37,6 х 106 |
Мышь, культура ткани | 10,83 | 5050 |
Человек, культура ткани | 11,24 | 3930 |
Крыса, гепатоцит | 16,1 | 3800 |
Мышь, лимфоцит | 3,3 | 400 |
Человек, лимфоцит | 4,47 | 700 |
По поверхности ядра поры располагаются более или менее равномерно, но их число резко падает в местах ассоциации с ядерной оболочкой участков гетерохроматина, ядрышкового организатора, теломерных участков.
Поровые комплексы могут встречаться и в других мембранных компонентах клетки, но гораздо реже, чем в ядерной оболочке. Иногда поровые комплексы видны в составе мембран гранулярного эндоплазматического ретикулума. Они обнаруживаются в составе окончатых мембран цитоплазмы, которые представляют собой тесно расположенные пачки замкнутых плоских мембранных мешков, сплошь пронизанных поровыми комплексами, имеющими такую же структуру, как и поры в ядерной оболочке.
Интересные данные были получены при морфометрическом изучении поровых комплексов в ядрах и окончатых мембранах бластодермы эмбрионов дрозофилы. Оказалось, что при переходе от синцитиальной к целлюлярной стадии, количество пор в оболочках ядер остается неизменным, а количество пор в окончатых пластинках вырастает примерно в 10 раз. В дальнейшем окончатые мембраны полностью исчезают. На основании этого было сделано предположение, что на ранней стадии развития в бластодерме дрозофилы происходит “суперпродукция” поровых комплексов (или их компонентов), избыток которых “встраивается” в окончатые мембраны. Т.е. макромолекулярный ансамбль, составляющий комплекс ядерных пор, способен к автономной самосборке и к последующему встраиванию в различные мембранные системы.
Роль ядерной оболочки в ядерно-цитоплазматическом обмене
Со времени открытия ядерной оболочки и описания ее строения делалось заключение о том, что ядерная оболочка может служить регулятором в ядерно-цитоплазматическом обмене и главная роль в этих процессах отводилась ядерным порам. Обмен же продуктами между ядром и цитоплазмой в самом деле очень велик: все ядерные белки поступают в ядро из цитоплазмы и все формы РНК выводятся из ядер. И в этом процессе комплекс поры выступает как супрамолекулярный комплекс, выполняющий роль не только транслокатора, механизма переноса, но и роль сортировщика, узнающего и отбирающего специальным образом переносимые молекулы.
В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некоторое молекулярное сито, пропуская частицы определенного размера пассивно, по градиенту концентрации. Так, ионы, сахара, нуклеотиды, АТФ, гормоны — свободно поступают в ядра. С другой стороны ядерные поры осуществляют избирательный транспорт.
Через ядерную оболочку беспрепятственно в обе стороны происходит пассивный транспорт высоко молекулярных соединений, имеющих массу не более 5 х 103 дальтон. Для определения размеров частиц, могущих пройти сквозь пору, используются гранулы декстрана или коллоидного золота, которые путем микроинъекции вводятся в цитоплазму живой клетки. Было обнаружено, что максимальный размер частиц, способных транспортироваться в ядро составляет 8,5-10 нм. При этом сначала частицы собираются в зоне поровых комплексов, а затем оказываются в ядре. Неядерные белки с массой большей, чем 20 000-40 000 дальтон проникают в ядро медленнее, если вообще проникают. Так инъецированные белки с массой 17 кД могут проникнуть в ядро довольно быстро, за 2-3 минуты, белки 40 кД — за 30 минут, белки 60 кД — вообще не проникают в ядра. Считается, что белки с гидродинамическим радиусом больше 3,5 нм (что соответствует глобулярному белку с массой 65 кД), не могут просто механически проходить через ядерную пору. В этих случаях ядерная пора выступает в качестве реального молекулярного сита.
Но дело осложняется тем, что многие белки поступают как в ядро, так и выходят из него против градиента концентраций. Так, например, концентрация гистонов в ядре значительно выше, чем в цитоплазме. Но, несмотря на это, во время синтеза ДНК происходит транспорт огромного количества (106 молекул каждые три минуты, или по 100-500 молекул через одну пору за 1 минуту) гистонов из цитоплазмы в ядро. С другой стороны через ядерные поры реально могут проходить некоторые белки и даже макромолекулярные комплексы с массой значительно большей, чем 60 кД.
Через ядерные поры из цитоплазмы в ядро транспортируются крупные молекулы белков, например, белок нуклеоплазмин, пентамер с молекулярной массой 125 кД. Из ядра через поры выходят в цитоплазму субъединицы рибосом и другие рибонуклеопротеиды, меньшие из которых могут иметь массу 250 кД. Эти данные показывают, что комплексы ядерных пор не представляют собой просто механические сита, которые ограничивают транспорт молекул в зависимости от их размеров.
Работы последнего времени показывают, что многие ядерные белки проходят через ядерные поры с помощью специальных механизмов, включающих узнавание и связывание крупных ядерных белков, а затем только их транслокацию, перенос через поры. Было найдено, что белки, транспортируемые в ядро, имеют определенные последовательности аминокислот — последовательности ядерной локализации (NLS), которые узнаются рецепторами ядерных пор. Такие NLS характерны для кариофильных белков, т.е. для белков ядерной локализации, которые синтезируются на рибосомах в цитоплазме, а затем транспортируются в ядро.
Импорт кариофильных белков
Впервые аминокислотные последовательности ядерной локализации были обнаружены на С-конце субъединиц молекулы нуклеоплазмина (ядерный белок, принимающий участие в структуризации хроматина). Эти эксперименты были проведены на бесклеточной системе, когда выделенные ядра помещались в цитоплазматический экстракт ооцитов ксенопуса, куда добавляли нативные или измененные молекулы нуклеоплазмина. Это крупный белок (125 кД), состоящий из пяти субъединиц, каждая из которых обладает глобулярной и фибриллярной, С-концевой, частями. Если удалит путем протеолиза примерно 50 аминокислот с С-конца, то ни пентамер, ни мономеры в ядро не попадают, в то время как отщепленные фибриллярные участки через поры проходят свободно, так как содержат NLS-участок.
Более того, при смешивании неядерных белков с этими NLS-фрагментами, такие комплексы способны транспортироваться в ядро. Даже крупные частицы декстрана (20 нм), неспособные проникать в ядро, при связывании с ними NLS-последовательностей нуклеоплазмина транспортировались из цитоплазматического экстракта в ядро.
Подробно строение NLS изучено у белка Т антигена вируса SV40. Кариофильный сигнал состоял из последовательности: Pro-Lys-128Lys-Lys-Arg-Lys-Val. Одна лишь аминокислотная замена (128Lys на Thr или Asp) полностью лишают этого фрагмента кариофильных свойств. Оказалось. что можно создавать химерные белки с этим аминокислотным доменом, что позволяет необычные для ядер белки (альбумин плазмы, иммуноглобулин G, и даже ферритин с мол. массой 465 кД) транспортировать через ядерные поры.
Было показано, что белок с NLS проходит в ядро через несколько этапов (рис. 111). Импорт начинается с того, что NLS-белок связывается с гетеродимером рецептора NLS, с белками импортинами и , локализованными в цитоплазме. Возникший белковый комплекс (импортируемый белок с NLS, связанный с импортинами и ) подходит к внешней ядерной мембране и закрепляется на цитоплазматических филаментах порового комплекса. Затем этот комплекс входит в ядерную пору и проходит через “транспортер”. Считается, что транспортер состоит из множества извитых белковых филаментов, обогащенных аланином и глицином (FG-филаменты), представляющих собой барьер для транспорта некариофильных белков. Комплекс же, имеющий NLS как бы разрыхляет эту сеть и проходит через канал транспортера. После перехода комплекса в нуклеоплазму импортин связывается с белком RAN, представляющего собой малую GTP-азу, что приводит к распаду комплекса. Импортируемый белок освобождается и остается в ядре, импортин возвращается в цитоплазму, так же как и импортин , но в связи с RAN-GDP, где последние также диссоциируют. Тем самым только белок с NLS остается в составе ядра (рис. 111).
Экспорт из ядра в цитоплазму
Из ядра в цитоплазму также существует поток как белков, так и ядерных транскриптов в виде рибонуклеопротеидов. В принципе этот экспорт своей организации сходен с процессом импорта кариофильных белков. Так было обнаружено, что гликопротеидные молекулы, связывающие лиганды, локализуются в место поровых комплексов и со стороны ядра. Одна и та же пора может принимать участие как в импорте, так и в экспорте макромолекул. В пользу этого говорит то, что частички коллоидного золота, связанного с нуклеоплазмином, сорбируются на ядерной поре со стороны цитоплазмы, одновременно с сорбцией частичек, связанных с РНК, инъецированных в ядре ооцитов. Подобные эксперименты показали, что многие РНК (тРНК, 5S РНК, поли-У и поли-А), связанные с коллоидным золотом, аккумулируются в зоне ядерных пор, а затем переносятся в цитоплазму. Более того, РНК способствует переносу через ядерную пору крупных частиц золота размером до 20 нм. Обратного переноса не происходит: аналогичные частички, инъецированные в цитоплазму ооцита, в ядро не проникают.
Что касается естественных видов РНП, то комплексы ядерных пор также должны узнавать специфический сигнал на экспорт. Белковые компоненты РНП несут аминокислотные последовательности, сигналы ядерного экспорта (NES), которые дают возможность различным РНП проходить через ядерную оболочку в цитоплазму.
В этом случае также образуется сложный комплекс, состоящий из переносимого белка с NES-последовательностью (связанного с РНК или свободного), ассоциированного с белком экспортином 1, который в свою очередь связан с RAN-GTP. Этот комплекс проходит через центральный канал, создаваемый транспортером, в цитоплазму, где и диссоциирует. При этом освобождается белок с NES-участком (или РНП), который остается в цитоплазме. Экспортин 1 и RAN после гидролиза GTP снова возвращаются в ядро (рис. 112).
В процессе экспорта РНП ядерная пора контролирует не только белковый компонент. Ядерные поры узнают и не экспортируют короткие (100 нуклеотидов) тРНК, если в их структуре есть хоть одна замена. Транспорт незрелых форм иРНК, имеющих интронные участки, не происходит. Вообще в цитоплазме не обнаруживаются незрелые РНК. Вероятно, для экспорта некоторых РНК необходима их связь с особыми белками. Так 5S РНК переносится в цитоплазму вместе с транскрипционным фактором TFIIIA, или с белком L5. Мутантные формы 5S рРНК, которые не связываются с TFIIIA, остаются в ядре.
Мало изучен вопрос о транспорте в цитоплазму крупных РНП-комплексов, таких как субъединицы рибосом, информосомы и малые ядерные РНП. Возможно все они под действием каких-то факторов разворачиваются, меняют свою конформацию и проходят через поровый комплекс. В пользу этого говорят наблюдения гантелевидных РНК-содержащих частиц, в просвете пор ядер гигантских слюнных желез насекомых. Считается, что эта картина отражает момент выхода из ядер РНП-частиц 60 нм в диаметре, относимых к информоферам. Интересно, что состав белков в цитоплазматических информационных РНП иной, это может говорить о том, что в зоне поровых комплексов происходит “переодевание” информационных РНК, связь их с иными белками.
Динамика ядерной оболочки в митозе
Большей частью, но не у всех видов (исключение составляют амебы, эвгленовые, инфузории, динофлагелляты, многие водоросли, некоторые грибы), ядерная оболочка разрушается при митозе и снова возникает после деления клеток. Это так называемый открытый тип митоза (рис. 113). При этом в профазе по мере конденсации хромосом ядерная оболочка теряет с ними связь, а затем в ней появляются разрывы. Она приобретает вид плоских мембранных вакуолей, цистерн. В это время ядерные поры еще видны. Позднее они исчезают. Во время митоза 120 мДа комплекс ядерной поры разбирается на субкомплексы примерно по 1 мДа. Разборка пор начинается с фосфорилирования ряда нуклеопоринов митотической cdc2/циклин B киназой.
Ядерная оболочка превращается в скопление мелких мембранных пузырьков, окружающих зону бывшего интерфазного ядра. Такие пузырьки морфологически нельзя отличить от других мелких вакуолей в цитоплазме, они вероятно сливаются с вакуолями эндоплазматического ретикулума. В метафазе мембранные элементы цитоплазмы оттесняются к периферическим зонам клеток микротрубочками веретена деления.
В конце анафазы, когда прекращается движение хромосом к противоположным полюсам клетки, мембранные пузырьки цитоплазмы, и в первую очередь мембраны гранулярного эндоплазматического ретикулума (см. ниже), начинают контактировать с поверхностью хромосом. Эти контакты происходят сначала в небольшом числе точек, но затем начинается перестройка и рост этих первичных зачатков ядерной оболочки. Они из мелких пузырьков превращаются в плоские вакуоли, которые растут в ширину и обволакивают поверхность деконденсирующихся хромосом. Участки таких растущих плоских мембранных мешков сливаются, замыкая и отгораживая содержимое нового интерфазного ядра. Интересно, что ядерные поры появляются на самых ранних этапах реконструкции ядерной оболочки, когда двойные мембранные цистерны еще не сомкнулись и фактически ничего не разделяют.
При реконструкции ядерной оболочки происходит сборка ядерных пор. Она начинается с образования ямки при слиянии внешней и внутренней ядерной мембраны, которая затем превращается в отверстие. В этом процессе принимают участие интегральные белки gp 210 и POM 121, которые впоследствии будут закреплять ЯПК на мембранах.
За этим следует появление внутренних структур ЯПК: комплекс кольца, спиц, добавление звездчатого кольца и других структур, и, наконец, филаментов.
У некоторых низших организмов в случае закрытого митоза ядерная оболочка не исчезает, она в зоне ядерной перетяжки замыкается, что приводит к образованию двух новых ядер. Здесь участие ядерной оболочки в делении клетки заключается в том, что на ней закреплены хромосомы, и она, по-видимому, принимает участие в индукции образования микротрубочек, необходимых при делении клеток.
По-видимому, для реконструкции ядерной оболочки необходимым условием является деконденсация хромосом. Было показано, что если вызвать преждевременную деконденсацию метафазных хромосом, то они очень быстро контактируют с мембранными пузырьками и одеваются каждая своей отдельно ядерной оболочкой, вследствие чего в клетке возникает множество так называемых микроядер, каждое их которых возникло из одной хромосомы.
С другой стороны, можно экспериментально вызвать разборку ядерной оболочки у интерфазного ядра. Это происходит, если слить в культуре ткани две клетки на разных стадиях клеточного цикла и получить т.н. гетерокарион, где одно из ядер будет находиться в интерфазе, а другое быть в виде митотических хромосом в метафазе. В этом случае в интерфазном ядре начинает конденсироваться хроматин, образуются преждевременно конденсированные хромосомы, а ядерная оболочка исчезнет так же как во время нормального митоза (рис. 114). Эти данные говорят о том, что в цитоплазме митотической клетки существуют какие-то факторы, вызывающие как конденсацию хромосом, так и параллельный этому процесс распада ядерной оболочки.
Сходная динамика совпадения процессов перестройки хромосом и ядерной оболочки наблюдается и в другой системе, в цитоплазме ооцитов или в бесклеточных цитоплазматических экстрактах ооцитов. Так если в цитоплазму ооцита амфибий на стадии метафазы инъецировать выделенные интерфазные ядра, то их ядерная оболочка разбирается, а хроматин конденсируется в виде митотических хромосом. Если же в ооцит на стадии интерфазы ввести митотические хромосомы, то они начинают деконденсироваться, появляются множественные мелкие вакуоли, которые сливаясь друг с другом, образуют ядерные оболочки. Интересно, что в цитоплазму интерфазного ооцита можно ввести даже чужеродную чистую ДНК, которая, связываясь с гистонами в цитоплазме, образует хроматиновые глыбки, которые в свою очередь одеваются ядерными оболочками и превращаются в микроядра.
Эти экспериментальные приемы вместе с методом иммунофлуоресценции позволили проследить судьбу многих белков ядерной оболочки во время митоза. Подробно изучена судьба ламинов. Было найдено, что фиброзный слой ламинов деполимеризуется параллельно распаду ядерных мембран и конденсации хроматина. Этому предшествует обильное (в 7 раз выше, чем в интерфазе) фосфорилирование ламинов. Ламины A и C при этом деполимеризуются до димеров и тетрамеров и, переходя в растворимое состояние, равномерно распределяются в цитоплазме вне связи с другими структурами. Ламин B тоже деполимеризуется до олигомеров, но остается связанным с мембранными пузырьками, возникшими из ядерной оболочки.
При сборке ядерной оболочки в телофазе белки ламины иммунохимически начинают выявляться в центромерных и теломерных участках хромосом, там же обнаруживаются первые признаки образования новой ядерной оболочки. Там же накапливаются антитела к белкам порового комплекса. В бесклеточной системе цитоплазматического экстракта ооцитов было показано, что ассоциация растворимых в митозе ламинов A и C происходит независимо от ламина B. Оказалось, что если систему реконструкции ядерной оболочки лишить ламина B, то ламины A и C связываются с поверхностью хромосом, но сборки ядерной оболочки не происходит. В экстракте, лишенном ламинов A и C, ламин B связывается с хромосомами, но нормальная ядерная оболочка так же не формируется.
studfiles.net
чем отличается мембрана клетки от ядерной оболочки и в чём их сходство?
Говоря о клеточном ядре, мы имеем в виду собственно ядра эукариотических клеток. Их ядра построены сложным образом и довольно резко отличаются от “ядерных” образований, нуклеоидов, прокариотических организмов. У последних в состав нуклеоидов (ядроподобных структур) входит одиночная кольцевая молекула ДНК, практически лишенная белков. Иногда такую молекулу ДНК бактериальных клеток называют бактериальной хромосомой, или генофором (носителем генов). Бактериальная хромосома не отделена мембранами от основной цитоплазмы, однако собрана в компактную ядерную зону — нуклеоид, который можно видеть в световом микроскопе после специальных окрасок. Сам термин “ядро” впервые был применен Броуном в 1833 г. Для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов. Клеточное ядро обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, кариоплазмы (или ядерного сока) (рис). Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариоти-ческих одно- и многоклеточных организмов. Ядра имеют обычно шаровидную или яйцевидную форму; диаметр первых равен приблизительно 10 мкм, а длина вторых — 20 мкм. Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю ее активность. Связано это с тем, что ядро несет в себе генетическую (наследственную) информацию, заключенную в ДНК. Ядерная оболочка Эта структура характерна для всех эукариотических клеток. Ядерная оболочка состоит из внешней и внутренней мембран, разделенных перинуклеарным пространством шириной от 20 до 60 нм. В состав ядерной оболочки входят ядерные поры. Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран: они имеют толщину около 7 нм и состоят из двух осмиофильных слоев. В общем виде ядерная оболочка может быть представлена, как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы. Из всех внутриклеточных мембранных компонентов таким типом расположения мембран обладают только ядро, митохондрии и пластиды. Однако ядерная оболочка имеет характерную особенность, отличающую ее от других мембранных структур клетки. Это наличие особых пор в оболочке ядра, которые образуются за счет многочисленных зон слияний двух ядерных мембран и представляет собой как бы округлые перфорации всей ядерной оболочки. Строение ядерной оболочки Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд сруктурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматического ретикулума. Так, на внешней ядерной мембране обычно располагается большое количество рибосом. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность — она может образовывать различной величины выпячивания или выросты в сторону цитоплазмы. Внутренняя мембрана контактирует с хромосомным материалом ядра (см. Ниже). Наиболее характерной и бросающейся в глаза структурой в ядерной оболочке является ядерная пора. Поры в оболочке образуются за счет слияния двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром 80-90 нм. Округлое сквозное отверстие в ядерной оболочке заполнено сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом пор ядра.
touch.otvet.mail.ru
особенности строения, функции ядра и значение для клетки
Ядро – главное составляющее живой клетки, которое несет наследственную информацию, закодированную набором генов. Оно занимает центральное положение в клетке. Размеры варьируются, форма обычно сферичная или овальная. В диаметре ядро в разных клетках может быть от 8 до 25мкм. Есть исключения, примеру, яйцеклетки рыб имеют ядра диаметром в 1 мм.
Особенности строения ядра
Заполнено ядро жидкостью и несколькими структурными элементами. В нем выделяют оболочку, набор хромосом, нуклеоплазму, ядрышка. Оболочка двухмембранная, между мембранами находится перенуклеарное пространство.
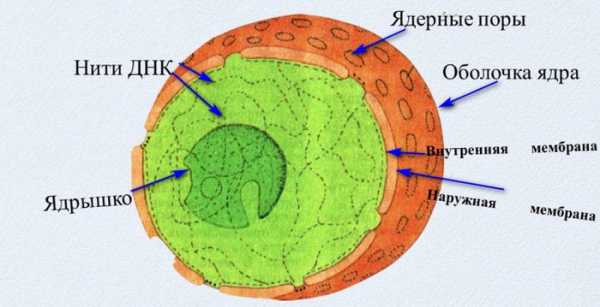
Внешняя мембрана сходна по строению с эндоплазматическим ретикулумом. Она связана с ЭПР, который будто ответвляется от ядерной оболочки. Снаружи на ядре находятся рибосомы.
Внутренняя мембрана прочная, так как в ее состав входит ламина. Она выполняет опорную функцию и служит местом крепления для хроматина.
Мембрана имеет поры, обеспечивающие обменные процессы с цитоплазмой. Ядерные поры состоят из транспортных белков, которые поставляют в кариоплазму вещества путем активного транспорта. Пассивно сквозь поровые отверстия могут пройти только небольшие молекулы. Также каждая пора прикрыта поросомой, которая регулирует обменные процессы в ядре.
Количество ядер в разных по специализации клетках различно. В большинстве случаев клетки одноядерные, но есть ткани, построенные из многоядерных клеток (печеночная или ткань мозга). Есть клетки лишенные ядра – это зрелые эритроциты.
У простейших выделяют два типа ядер: одни отвечают за сохранение информации, другие – за синтез белка.
Ядро может прибывать в состоянии покоя (период интерфазы) или деления. Переходя в интерфазу, имеет вид сферического образования с множеством гранул белого цвета (хроматина). Хроматин бывает двух видов: гетерохроматин и эухроматин.
Эухроматин – это активный хроматин, который сохраняет деспирализированное строение в покоящемся ядре, способен к интенсивному синтезу РНК.
Гетерохроматин – это участки хроматина, которые находятся в конденсированном состоянии. Он может при необходимости переходить в эухроматиновое состояние.
При использовании цитологического метода окрашивания ядра (по Романовскому-Гимзе) выявлено, что гетерохроматин меняет цвет, а эухроматин нет. Хроматин построен из нуклеопротеидных нитей, названных хромосомами. Хромосомы несут в себе основную генетическую информацию каждого человека. Хроматин — форма существования наследственной информации в интерфазном периоде клеточного цикла, во время деления он трансформируется в хромосомы.
Строение хромосом
Каждая хромосома построена из пары хроматид, которые находятся параллельно друг к другу и связаны только в одном месте – центромере. Центромера разделяет хромосому на два плеча. В зависимости от длины плеч выделяют три вида хромосом:
- Равноплечие;
- разноплечие,
- одноплечие.

Некоторые хромосомы имеют дополнительный участок, который крепится к основному нитевидными соединениями – это сателлит. Сателлиты помогают идентифицировать разные пары хромосом.
Метафазное ядро представляет собой пластинку, где располагаются хромосомы. Именно в эту фазу митоза изучается количество и строение хромосом. Во время метафазы сестринские хромосомы двигаются в центр и распадаются на две хроматиды.
Строение ядрышка
В ядре также находится немембранное образование — ядрышко. Ядрышки представляют собой уплотненные, округлые тельца, способные преломлять свет. Это основное место синтеза рибосомальной РНК и необходимых белков.
Число ядрышек различно в разных клетках, они могут объединяться в одно крупное образование или существовать отдельно друг от друга в виде мелких частиц. При активации синтетических процессов объем ядрышка увеличивается. Оно лишено оболочки и находится в окружении конденсированного хроматина. В ядрышке также содержатся металлы, в большей мере цинк. Таким образом, ядрышко – это динамичное, меняющееся образование, необходимое для синтеза РНК и транспорта ее в цитоплазму.
Нуклеоплазма заполняет все внутреннее пространство ядра. В нуклеоплазме находится ДНК, РНК, протеиновые молекулы, ферментативные вещества.
Функции ядра в клетке
- Принимает участие в синтезе белка, рибосомной РНК.
- Регулирует функциональную активность клетки.
- Сохранение генетической информации, точная ее репликация и передача потомству.
Роль и значение ядра
Ядро является главным хранилищем наследственной информации и определяет фенотип организма. В ядре ДНК существует в неизмененном виде благодаря репарационным ядерным ферментам, которые способны ликвидировать поломки и мутации. Во время клеточного деления ядерные механизмы обеспечивают точное и равномерное расхождение генетической информации в дочерние клетки.
animals-world.ru
строение мембраны, участие в процессе деления
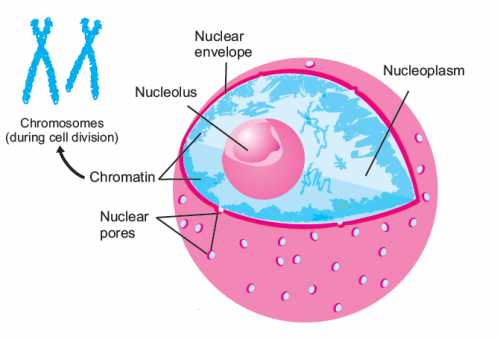
Ядерная оболочка (нуклеолемма) представляет собой сложное образование, отделяющее содержимое ядра от цитоплазмы и других элементов живой клетки. Данная оболочка выполняет ряд важных функций, без которых является невозможным функционирование ядер, полноценное деление клеток. Чтобы определить роль ядерных мембран в жизнедеятельности эукариотных клеток, необходимо узнать не только главные функции, но и особенности строения.
В статье подробно рассматриваются функции ядерной оболочки. Описывается строение, структурные компоненты нуклеолеммы, их взаимосвязь, механизмы транспортировки веществ, процесс деления при митозе.
Содержание:
Строение оболочки
Строение и свойства ЯПК
Виды ядерного транспорта
Импорт и экспорт белков
Ядерная оболочка при делении
Строение оболочки
Главное отличие эукариот заключается в наличии ядра и ряда других органелл, необходимых для его поддержания. Такие клетки входят в состав всех растений, грибов, животных, в то время как клетки-прокариоты представляют собой простейшие безъядерные организмы.
Нуклеолемма состоит из двух структурных элементов – внутренней и наружной мембран. В промежутке между ними существует свободное пространство, называемое перинуклеарным. Ширина перинуклеарного промежутка нуклеолеммы составляет от 20 до 60 нанометров (нм).
Внешняя мембрана нуклеолеммы контактирует с клеточной цитоплазмой. На ее наружной поверхности располагается существенное число рибосом, которые отвечают за синтез белковых соединений из отдельных аминокислот. Внешняя мембрана не содержит рибосом.

Мембраны, образующие нуклеолемму, состоят из белковых соединений и двойного слоя фосфолипидных веществ. Механическая прочность оболочки обеспечивается сетью филаментов – нитевидных белковых структур. Наличие филаментной сети характерно для большинства эукариот. Они соприкасаются с внутренней мембраной.
Сети филаментов располагаются не только в области нукелолеммах. Такие структуры также располагаются в цитоплазме. Их функция заключается в сохранении целостности клетки, а также в формировании контактов между клетками. При этом, отмечается, что слои, образующие сеть, регулярно перестраиваются. Данный процесс наиболее активен в период роста клеточного ядра перед делением.
Сеть филаментов, которая поддерживает мембраны, называется ядерной ламиной. Она формируется из определенной последовательности белков-полимеров, которые называются ламинами. Она взаимодействует с хроматином – веществом, участвующим в формировании хромосом. Также ламина контактирует с молекулами рибонуклеиновой кислоты, ответственными за хранение генетической информации.
Внешняя мембрана ядра взаимодействует с мембраной, окружающей эндоплазматический ретикулум. В определенных участках оболочки происходит контакт перинуклеарного пространства и внутреннего пространства ретикулума.
Функции эндоплазматического ретикулума:
- Синтез и транспортировка белков
- Хранение продуктов синтеза
- Формирование новой оболочки при митозе
- Хранение минералов кальция, выполняющих функцию медиатора
- Продукция гормонов
Внутри оболочки располагаются ядерные поровые комплексы. Это каналы, посредством которых происходит перенос молекул между клеточным ядром, цитоплазмой и другими клеточными органеллами. На одном квадратном микроне поверхности нуклеолеммы располагает от 10 до 20 поровых комплекса. Исходя из этого, в оболочке 1 соматической клетки может находится всего от 2 до 4 тысяч ЯПК.
Помимо транспорта веществ, оболочка выполняет опорную и защитную функцию. Она отделяет ядро от содержимого цитоплазмы, в том числе продуктов деятельности других органелл. Защитная функция заключается в предохранении генетической информации ядра от негативного воздействия, например, вирусной инфекции.
Считается, что двойная мембрана ядерной оболочки сформировалась в ходе эволюции путем захвата одних клеток другими. Вследствие этого, некоторые поглощенные клетки сохранили собственную активность, но при этом их ядро было окружено двойной мембраной – собственной, и мембраной клетки-хозяина.
Таким образом, ядерная оболочка представляет собой сложную структуру, состоящую из двойной мембраны, содержащей ядерные поры.
Строение и свойства ЯПК
Ядерный поровый комплекс – это симметричный канал, местом локализации которого является сличение наружной и внутренней мембран. ЯПК состоят из набора веществ, включающих около 30 видов белков.
Ядерные поры имеют бочкообразную форму. Образуемый канал не ограничивается ядерными мембранами, а незначительно выступает за их пределы. В результате с двух сторон оболочки возникают кольцеобразные выступы. Размер этих выступов отличается, так как с одной стороны кольцеобразное образование имеет больший диаметр, чем с другой. Элементы ядерных пор, выступающих за пределы мембраны, называются терминальными структурами.
Цитоплазматическая терминальная структура (та что находится на внешней поверхности ядерной мембраны) состоит из восьми коротких фибрилл-нитей. Ядерная терминальная структура также состоит из 8 фибрилл, однако они образуют кольцо, выполняющего функции корзины. Во многих клетках от ядерной корзины исходят дополнительные фибриллы. Терминальные структуры являются местами, где происходит контакт молекул, транспортируемых через ядерные поры.
В месте расположения ЯПК происходит слияние наружной и внутренней ядерной мембраны. Такое слияние объясняется необходимостью обеспечить фиксацию ядерных пор в мембранах с помощью белков, соединяющих их также с ядерной ламиной.
В настоящее время общепринятым считается модульное строение ядерных каналов. Такая модель предусматривает структуру поры, состоящую из нескольких кольцевидных образований.
Внутри ядерной поры постоянно находится плотное вещество. Ее происхождение точно не известно, однако считается, что оно является одним из элементов ЯПК, за счет которого осуществляется транспортировка молекул от цитоплазмы к ядру и наоборот. Благодаря исследованию с использованием электронных микроскопов с высоким разрешением удалось выяснить, что плотная среда внутри ядерного канала способна менять свое месторасположение. Ввиду этого, считается, плотная внутренняя среда ЯПК является карго-рецепторным комплексом.
Транспортные функции ядерной оболочки возможны благодаря наличию ядерных поровых комплексов.
Виды ядерного транспорта
Транспортировка веществ через ядерную оболочку называется ядерно-цитоплазматическим транспортом веществ. Данный процесс предусматривает своеобразный обмен молекулами, синтезируемыми в ядре, и веществами, обеспечивающими жизнедеятельность самого ядра, импортируемыми из цитоплазмы.
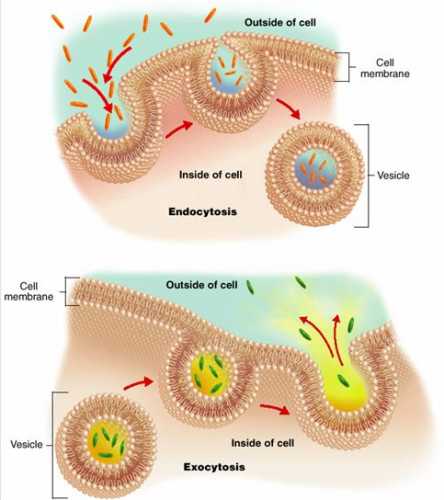
Существуют такие виды транспортировки:
- Пассивная. Посредством данного процесса осуществляется перемещение небольших молекул. В частности, через пассивный транспорт происходит передача мононуклеотидов, минеральных компонентов, продуктов метаболического обмена. Процесс называется пассивным, так как протекает путем диффузии. Скорость прохождения через ядерную пору зависит от размера вещества. Чем оно меньше, тем выше скорость транспортировки.
- Активная. Предусматривает перенос через каналы внутри ядерной оболочки крупных молекул или их соединений. При этом, соединения не распадаются на мелкие частицы, что позволило бы увеличить скорость транспортировки. Данный процесс обеспечивает поступление в цитоплазму синтезируемых в ядре рибонуклеиновых молекул. Из внешнего цитоплазматического пространства за счет активного транспорта происходит перенос белков, необходимых для метаболических процессов.
Выделяют пассивный и активный транспорт белков, отличающийся механизмом действия.
Импорт и экспорт белков
Рассматривая функции ядерной оболочки, необходимо напомнить о том, что транспортировка веществ осуществляется в двух направлениях – из цитоплазмы в ядро и наоборот.
Импорт белковых соединений через мембраны к ядру осуществляется за счет наличия особых рецепторов, называемых транспортинами. Эти компоненты содержат запрограммированный сигнал, за счет которого происходит движение в необходимом направлении. Белки и соединения, не обладающие таким сигналом, способны присоединятся к веществам, у которых он есть, и таким образом беспрепятственно перемещаться.
Важно отметить, что сигналы ядерного импорта обеспечивают избирательность поступления веществ в ядро. Многие образования, в числе которых полимеразы ДНК и РНК, а также белки, участвующие в процессах регуляции, не достигают ядра. Таким образом, ядерные поры представляют собой не только механизм транспортировки веществ, но и их своеобразной сортировки.

Сигнальные белки отличаются друг от друга. Ввиду этого, существует разница между скоростью перемещения через поры. Также они выполняют функцию источника энергии, так как для перемещения крупных молекул, транспортировка которых не возможна диффузным путем, необходимы дополнительные энергетические затраты.
Первый этап импорта белков заключается в присоединении к импортину (транспортину, обеспечивающему перенос через канал к ядру). Полученное в результате слияния сложное образование проходит через ядерную пору. После этого, с ним связывается другое вещество, за счет которого транспортируемый белок освобождается, а импортин возвращается обратно в цитоплазму. Таким образом, импорт в ядро представляет собой цикличный замкнутый процесс.
Транспорт веществ из ядра через оболочку в цитоплазматическое пространство осуществляется аналогичным образом. Исключением является то, что за перенос вещества-груза отвечают уже сигнальные белки, называемые экспортинами.
На первой стадии процесса белок (в большинстве случаев это молекулы РНК) связываются с экспортином и веществом, отвечающим за высвобождение транспортируемого субстрата. После перехода сквозь оболочку нуклеотид расщепляется, за счет чего переносимый белок высвобождается.
В целом, перенос веществ между ядром и цитоплазмой представляет собой цикличный процесс, осуществляемый за счет белков-транспортинов и веществ, отвечающих за высвобождение груза.
Ядерная оболочка при делении
Большинство клеток-эукариотов размножаются путем непрямого деления, которое называется митозом. Данный процесс предусматривает разделение ядра и других клеточных структур с сохранением одинакового количества хромосом. За счет этого сохраняется генетическая идентичность, полученная в результате деления клеток.
В процессе деления нуклеолемма выполняет еще одну важную функцию. После того как происходит разрушение ядра, внутренняя мембрана не позволяет хромосомам расходится на большие расстояния друг от друга. Хромосомы фиксируются на поверхности мембраны до момента полноценного деления ядер и формирования новой нуклеолеммы.
Ядерная оболочка, несомненно, принимает активное участие в клеточном делении. Процесс состоит из двух последовательных этапов – разрушения и перестройки.
Распад ядерной оболочки происходит в прометафазе. Разрушение мембран происходит стремительно. После распада хромосомы характеризуются хаотичным расположением в области ранее существовавшего ядра. В дальнейшем образуется веретено деления – биполярная структура, между полюсами которой формируются микротрубочки. Веретено обеспечивает деление хромосом и их распределение между двумя дочерними клетками.
Перераспределение хромосом и формирование новых ядерных мембран происходит в период телофазы. Точный механизм восстановления оболочек не известен. Распространенной является теория о том, что слияние частиц разрушенной оболочки происходит под действием везикул – мелких клеточных органелл, функция которых заключается в сборе и хранении питательных веществ.
Также образование новых ядерных мембран связывают с переформировкой эндоплазматического ретикулума. Из разрушенного ЭПР высвобождаются белковые соединения, которые постепенно обволакивают пространство вокруг нового ядра, в результате чего в дальнейшем образуется целостная мембранная поверхность.
Таким образом, нуклеолемма принимает непосредственное участие в процессе деления клетки путем митоза.
Ядерная оболочка – сложный структурный компонент клетки, выполняющий барьерные, защитные, транспортные функции. Полноценное функционирование нуклеолеммы обеспечивается особенностями ее строения, взаимодействием с другими клеточными компонентами и биохимическими процессами, протекающими в них.
plushealth.ru
Ядро. Хромосомы. Митохондрии и Пластиды. Двухмембранные структуры
Двухмембранные структуры. Ядро. Хромосомы. Митохондрии и Пластиды
Ядро. Хромосомы. Митохондрии и Пластиды. Двухмембранные структуры
Ядро
Является непременным компонентом почти для каждой клетки эукариот (за исключением эритроцитов, тромбоцитов млекопитающих, ситовидных трубок растений). Клетки, как правило, имеют одно ядро, но встречаются двухядерные (инфузории) и многоядерные (гепатоциты, мышечные клетки и т. п.). Каждый тип клетки имеет определенное постоянное соотношение между объемами ядра и цитоплазмы – ядерно-цитоплазматическое соотношение.
Форма ядра
Ядра бывают разной формы и размеров. Обычная форма ядра – шарообразная, реже – другая (звездчатая, неправильная и т. п.). Размеры колеблются от 1 мкм до 1 см.
Некоторые одноклеточные (инфузории и т. п.) имеют два ядра: вегетативное и генеративное. Генеративное обеспечивает передачу генетической информации, вегетативное – регулирует синтез белков.
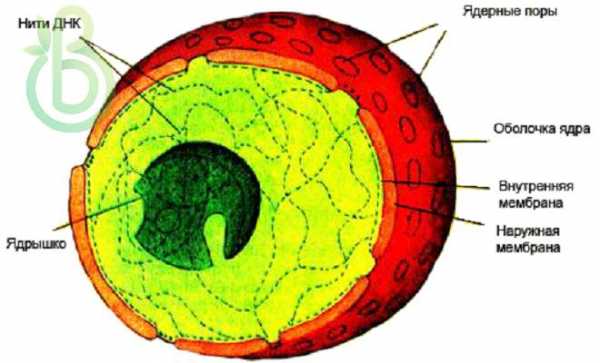
Покрыто двумя мембранами (внешней и внутренней) с ядерными порами, прикрытыми особыми тельцами; внутри – ядерный матрикс, состоящий из ядерного сока (кариоплазмы, нуклеоплазмы), ядрышек (одного или нескольких), рибонуклеопротеидных комплексов и нитей хроматина. Между двумя мембранами есть щель (от 20 до 60 нм). Внешняя мембрана ядра связана с ЭПС.
Внутреннее содержимое ядра
Кариоплазма (от греч. карион – ядро ореха) – это внутреннее содержимое ядра. По строению напоминает цитоплазму. Содержит белковые фибриллы, образующие внутренний скелет ядра.
Ядрышко состоит из комплекса РНК с белками (рибонуклеопротеидных фибрилл), внутреннеядрышкового хроматина и из предшественников субъединиц рибосом (гранул). Образуются на вторичных перетяжках хромосом – ядрышковых организаторах.
Функция ядрышек
Функция ядрышек: синтез рибосом.
Нити хроматина – хромосомы в период между делениями клетки (дезоксирибонуклеиновые комплексы). Имеют вид одиночных нитей (эухроматин), гранул (гетерохроматин) и интенсивно окрашиваются некоторыми красителями.
Хромосомы – ядерные структуры, в которых находятся гены, состоят из ДНК и белка. Кроме того, в состав хромосом входят ферменты и РНК.
Функции ядра
Сохранение и передача генетической информации, организация и регуляция процессов обмена веществ, физиологических и морфологических в клетке (например, синтез белка).
Хромосомы

Строение Хромосомы. Двухмембранные структуры
Хромосомы (от греч. хрома – цвет, сома – тело). Были открыты с помощью светового микроскопа в конце XIX века. Их строение лучше всего изучать на стадии метафазы митоза, когда они максимально спирализованы. Для этого располагают хромосомы по размерам (первые – наиболее длинные, последние – половые), составляют идеограммы.
Химический состав хромосом
В химический состав хромосом входят двухцепочечная ДНК, связанная с ядерными белками (образует нуклеопротеиды), РНК и ферменты. Ядерные белки, обернутые нитью ДНК, образуют нуклеосомы. По 8-10 нуклеосом соединяются в глобулы. Между ними содержатся участки ДНК. Таким образом компактно размещены в хромосоме молекулы ДНК. В развернутом виде молекулы ДНК очень длинные.
Состоят хромосомы из двух хроматид, соединенных первичной перетяжкой, которая разделяет их на плечи. Хромосомы могут быть равноплечими, разноплечими, одноплечими. В участке первичной перетяжки содержится пластинчатое образование в виде диска – центромера, к которому во время деления прикрепляются нити веретена деления. Могут иметь вторичную перетяжку (ядрышковый организатор) и спутник.
Каждая хромосома в наборе имеет себе подобную по строению и набору генов – гомологичную. Хромосомы разных пар будут по отношению одна к другой негомологичными. Хромосомы, которые не определяют пола, называются аутосомами. Хромосомы, определяющие пол, называются гетерохромосомами.
Какие бывают клетки
Клетки бывают неполовые – соматические (от греч. сома – тело) и половые, или генеративные (от лат. genero – порождаю, вырабатываю) гаметы. Количество хромосом в ядре может быть разным у разных видов организмов. Во всех соматических клетках организмов одного вида количество хромосом обычно одинаково. Для соматических характерен двойной набор хромосом – диплоидный (2n), для гамет – гаплоидный (n). Число хромосом может превышать двойной набор. Такой набор называется полиплоидным (триплоидный (Зn), тетраплоидный (4n) и т. п.).
Кариотип – это определенный набор хромосом в клетке, характерный для каждого вида растений, животных, грибов. Количество хромосом в кариотипе всегда четное. Количество хромосом не зависит от уровня организации организма, не всегда свидетельствует о филогенетическом родстве (у человека– 46 хромосом, у собаки – 78, у таракана – 48, у шимпанзе – 48).
Митохондрии
Митохондрии (от греч. митос – нить, хондрион – зернышко) – двухмембранные органеллы, которые имеют бобообразную форму палочек, нитей, есть почти во всех клетках эукариот. Иногда могут разветвляться (у некоторых одноклеточных, мышечных волокон и т. п.). Количество разное (от 1 до 100 тыс. и более). В клетках растений – меньше, поскольку их функцию (образование АТФ) частично выполняют хлоропласты.
Строение Митохондрии
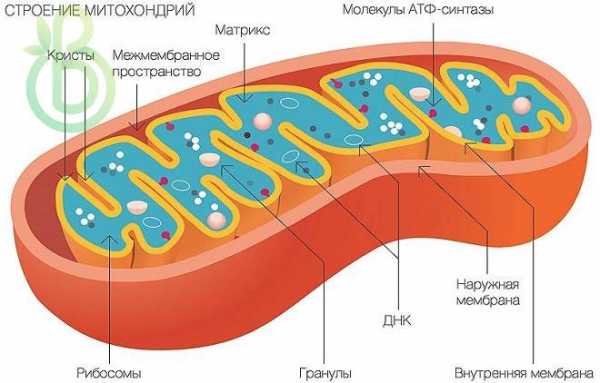
Строение Митохондрии. Двухмембранные структуры
Внешняя мембрана – гладкая, внутренняя – складчатая. Складки увеличивают внутреннюю поверхность, они называются кристами. Между внешней и внутренней мембранами есть щель (10-20 нм шириной). На поверхности внутренней мембраны расположен комплекс ферментов.
Внутренняя среда – матрикс. В нем находятся кольцевая молекула ДНК, рибосомы, иРНК, включения, синтезируются белки, входящие в состав внутренней мембраны.
Митохондрии в клетке постоянно восстанавливаются. Являются полуавтономными структурами – образуются путем деления.
Функции Митохондрии
Функции: энергетические «станции» клетки – образуют энергетически богатые вещества – АТФ, обеспечивают клеточное дыхание.
Пластиды
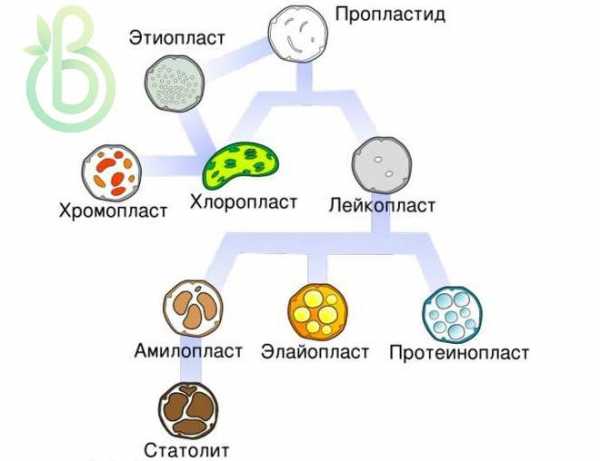
Пластиды. Двухмембранные структуры
Пластиды (от греч. пластидис, пластос – сформированный, вылепленный) – двухмембранные органеллы фотосинтезирующих организмов (преимущественно растений). Имеют разную форму, окраску. Различают три вида:
- Хлоропласты (от греч. хлорос – зеленый) – содержат в мембранах в основном хлорофилл, определяют зеленый цвет растений, находятся в зеленых частях растений. Длиной 5-10 мкм. Количество колеблется.
Строение хлоропластов
Строение: внешняя мембрана гладкая, внутренняя – складчатая, внутреннее содержимое – матрикс с кольцевой молекулой ДНК, рибосомами и включениями. Между внешней и внутренней мембранами – щель (20-30 нм). Внутренние мембраны образуют стопки – граны, которые состоят из тилакоидов (по 50 и больше), которые имеют вид уплощенных вакуолей или мешочков. Гран в хлоропласте 60 и более. Граны соединены ламеллами – плоскими удлиненными складками мембраны. На внутренних мембранах находятся фотосинтезирующие пигменты (хлорофилл и др.). Внутри хлоропласта – матрикс. В нем содержатся кольцевая молекула ДНК, рибосомы, включения, зерна крахмала.
Основные фотосинтезирующие пигменты (хлорофиллы, вспомогательные – каротиноиды) находятся в тилакоидах.
Основная функция хлоропластов
Основная функция – фотосинтез. В хлоропластах синтезируются также некоторые липиды, белки мембран.
Хлоропласты – полуавтономные структуры, располагают собственной генетической информацией, имеют собственный белоксинтезирующий аппарат, размножаются делением.
- Хромопласты (от греч. хрома – краска, цвет) – содержат цветные пигменты (каротины, ксантофиллы и др.), имеют немногочисленные тилакоиды, почти отсутствующую внутреннюю мембранную систему, находятся в цветных частях растения. Функции привлекают насекомых, других животных для опыления, рас-пространения плодов и семян.
- Лейкопласты (от греч. лейкос – белый) – это бесцветные пластиды, находятся в неокрашенных частях растения. Функция: запасают питательные вещества, продукты метаболизма клетки. Содержат кольцевую ДНК, рибосомы, включения, ферменты. Могут быть почти полностью заполнены зернами крахмала.
Пластиды имеют общее происхождение, возникают из пропластид образовательной ткани. Разные виды пластид могут переходить одна в другую. Светлые пропластиды превращаются в хлоропласты, лейкопласты –в хлоропласты или хромопласты. Разрушение хлорофилла в пластидах приводит к образованию хромопластов (осенью зеленая листва становится желтой, красной). Хромопласты – конечное преобразование пластид. Больше они ни в какие другие не превращаются.
У водорослей и некоторых жгутиковых есть особая двухмембранная органелла, которая содержит фотосинтезирующие пигменты – хроматофор. Она сходна по строению с хлоропластами, но имеет определенные отличия. В хроматофорах нет гран. Форма – разнообразная (у хламидомонады – чашевидная, у спирогиры – в виде спиральных лент и т. п.). В состав хроматофора входит пиреноид – участок клетки с мелкими вакуолями и зернами крахмала.
Гипотеза симбиогенеза (эндосимбиоза)
Клетки прокариот вступили в симбиоз с эукариотическими клетками. Считается, что митохондрии образовались в результате сожительства клеток аэробных и анаэробных, хлоропласты – в результате сожительства цианобактерий с клетками гетеротрофных первоначальных эукариот. Об этом свидетельствует то, что пластиды и митохондрии по размерам приближены к клеткам прокариот, имеют собственную кольцевую молекулу ДНК и собственный белоксинте- зирующий аппарат. Они являются полуавтономными, образуются путем деления.
xn—-9sbecybtxb6o.xn--p1ai
Ядерная мембрана — Википедия (с комментариями)
Материал из Википедии — свободной энциклопедии
Я́дерная мембра́на, или Я́дерная оболо́чка, или Кариоле́мма, или Кариоте́ка[1], или Нуклеоле́мма[2] — двойной липидный бислой, мембрана, окружающая ядро в эукариотических клетках.
Ядерная мембрана состоит из двух липидных бислоёв — наружной ядерной мембраны и внутренней ядерной мембраны. Пространство между мембранами называется перинуклеарным пространством; оно составляет единый компартмент с полостью с эндоплазматического ретикулума (ЭПР). Обычно ширина перинуклеарного пространства составляет около 20—40 нм[3].
Наружная мембрана
Наружная ядерная мембрана непосредственно переходит в мембрану эндоплазматического ретикулума[4], но при этом наружная ядерная мембрана содержит различные белки в значительно более высоких концентрациях, чем они присутствуют в ЭПР[5].
Внутренняя мембрана
Внутренняя мембрана ограничивает кариоплазму и изнутри покрыта ядерной ламиной, сетью промежуточных филаментов, которая поддерживает форму ядерной мембраны, обеспечивает прикрепление хроматина к оболочке ядра и участвует в регуляции экспрессии генов[5]. Ядерная ламина состоит из белков ламинов. Внутренняя мембрана связана с наружной мембраной посредством ядерных пор, пронизывающих обе мембраны. Хотя ЭПР и обе мембраны соединены друг с другом, многие белки, входящие в их состав, фиксированы в мембране, а не диффундируют свободно в её пределах[6].
Ядерные поры
Ядерная мембрана пронизана многочисленными ядерными порами. Это круппные белковые комплексы диаметром около 100 нм, с внутренней полостью около 40 нм шириной[5]. Они соединяют внутреннюю и наружную ядерные мембраны. Количество ядерных пор различно в разных типах клеток и может изменяться в зависимости от транскрипционной активности ядра.
Деление клетки
В течение G2-фазы интерфазы поверхность ядерной мембраны увеличивается, число ядерных пор иногда возрастает вдвое[5].
У некоторых низших эукариот, например, дрожжей, имеющих закрытый митоз, клеточная мембрана остаётся целой в ходе клеточного деления. Веретено деления у них формируется под мембраной[5]. При полузакрытом митозе в ядерной оболочке образуются крупные отверстия. При закрытом митозе с внеядерным веретеном (у динофлагеллят) в ядерную оболочку встраиваются центромеры хромосом.
У высших эукариот — животных и растений — ядерная мембрана разрушается в прометафазе митоза, позволяя веретену деления сформироваться снаружи. Механизм разрушения и перестройки ядерной мембраны ещё не до конца понятен.
Разрушение
У млекопитающих ядерная мембрана разрушается последовательно, шаг за шагом.
Сначала полипептиды нуклеопорины избирательно переносятся из ядерной мембраны. После этого оставшиеся ядерные поровые комплексы одновременно разрушаются. Биохимические исследования показали, что, скорее, ядерные поры распадаются на стабильные фрагменты, чем на короткие полипептидные цепочки[5].
Электронная и флуоресцентная микроскопия засвидетельствовали то, что ядерная мембрана абсорбируется эндоплазматическим ретикулумом — в норме ядерные белки в ЭПР не обнаруживаются, однако проявляются там в ходе митоза[5].
Перестройка
То, как ядерная мембрана вновь перестраивается в целостную структуру в течение телофазы, остаётся спорным вопросом. Существуют две теории[5]:
- Слияние везикул — везикулы ядерной мембраны сливаются, образуя ядерную мембрану;
- Переформировка ЭПР — части ЭПР, содержащие абсорбированные белки ядерной мембраны, покрывают ядерное пространство, формируя закрытую ядерную мембрану.
Напишите отзыв о статье «Ядерная мембрана»
Примечания
- ↑ [www.merriam-webster.com/dictionary/nuclear%20membrane nuclear membrane]. Merriam Webster. Проверено 7 декабря 2012. [www.webcitation.org/6FJKNWZh0 Архивировано из первоисточника 22 марта 2013].
- ↑ [www.biology-online.org/dictionary/Nuclear_membrane Nuclear membrane]. Biology Dictionary. Biology Online. Проверено 7 декабря 2012. [www.webcitation.org/6FJKOalrV Архивировано из первоисточника 22 марта 2013].
- ↑ [www.biology-online.org/dictionary/Perinuclear_space Perinuclear space]. Dictionary. Biology Online. Проверено 7 декабря 2012. [www.webcitation.org/6FJKPE99C Архивировано из первоисточника 22 марта 2013].
- ↑ [clapham.tch.harvard.edu/publications/pdf/Tabares_JMB_91.pdf Chloride channels in the Nuclear membrane]. Harvard.edu. Проверено 7 декабря 2012. [www.webcitation.org/6FJKPkm7A Архивировано из первоисточника 22 марта 2013].
- ↑ 1 2 3 4 5 6 7 8 Hetzer, Mertin (March 2, 2012). «[www.ncbi.nlm.nih.gov/pmc/articles/PMC2829960/ The Nuclear Envelope]». National Center for Biotechnology Information. Проверено 7 December 2012.
- ↑ (April 19, 2001) «[www.nature.com/emboj/journal/v20/n12/full/7593796a.html The inner nuclear membrane: simple, or very complex?]». The EMBO Journal. Проверено 7 December 2012.
Ссылки
- [sspatel.googlepages.com/nuclearporecomplex Animations of nuclear pores and transport through the nuclear envelope]
- [sspatel.googlepages.com/nuclearporecomplex2 Illustrations of nuclear pores and transport through the nuclear membrane]
- MeSH Nuclear+membrane
Отрывок, характеризующий Ядерная мембрана
Петя очнулся.– Уж светает, право, светает! – вскрикнул он.
Невидные прежде лошади стали видны до хвостов, и сквозь оголенные ветки виднелся водянистый свет. Петя встряхнулся, вскочил, достал из кармана целковый и дал Лихачеву, махнув, попробовал шашку и положил ее в ножны. Казаки отвязывали лошадей и подтягивали подпруги.
– Вот и командир, – сказал Лихачев. Из караулки вышел Денисов и, окликнув Петю, приказал собираться.
Быстро в полутьме разобрали лошадей, подтянули подпруги и разобрались по командам. Денисов стоял у караулки, отдавая последние приказания. Пехота партии, шлепая сотней ног, прошла вперед по дороге и быстро скрылась между деревьев в предрассветном тумане. Эсаул что то приказывал казакам. Петя держал свою лошадь в поводу, с нетерпением ожидая приказания садиться. Обмытое холодной водой, лицо его, в особенности глаза горели огнем, озноб пробегал по спине, и во всем теле что то быстро и равномерно дрожало.
– Ну, готово у вас все? – сказал Денисов. – Давай лошадей.
Лошадей подали. Денисов рассердился на казака за то, что подпруги были слабы, и, разбранив его, сел. Петя взялся за стремя. Лошадь, по привычке, хотела куснуть его за ногу, но Петя, не чувствуя своей тяжести, быстро вскочил в седло и, оглядываясь на тронувшихся сзади в темноте гусар, подъехал к Денисову.
– Василий Федорович, вы мне поручите что нибудь? Пожалуйста… ради бога… – сказал он. Денисов, казалось, забыл про существование Пети. Он оглянулся на него.
– Об одном тебя пг’ошу, – сказал он строго, – слушаться меня и никуда не соваться.
Во все время переезда Денисов ни слова не говорил больше с Петей и ехал молча. Когда подъехали к опушке леса, в поле заметно уже стало светлеть. Денисов поговорил что то шепотом с эсаулом, и казаки стали проезжать мимо Пети и Денисова. Когда они все проехали, Денисов тронул свою лошадь и поехал под гору. Садясь на зады и скользя, лошади спускались с своими седоками в лощину. Петя ехал рядом с Денисовым. Дрожь во всем его теле все усиливалась. Становилось все светлее и светлее, только туман скрывал отдаленные предметы. Съехав вниз и оглянувшись назад, Денисов кивнул головой казаку, стоявшему подле него.
– Сигнал! – проговорил он.
Казак поднял руку, раздался выстрел. И в то же мгновение послышался топот впереди поскакавших лошадей, крики с разных сторон и еще выстрелы.
В то же мгновение, как раздались первые звуки топота и крика, Петя, ударив свою лошадь и выпустив поводья, не слушая Денисова, кричавшего на него, поскакал вперед. Пете показалось, что вдруг совершенно, как середь дня, ярко рассвело в ту минуту, как послышался выстрел. Он подскакал к мосту. Впереди по дороге скакали казаки. На мосту он столкнулся с отставшим казаком и поскакал дальше. Впереди какие то люди, – должно быть, это были французы, – бежали с правой стороны дороги на левую. Один упал в грязь под ногами Петиной лошади.
У одной избы столпились казаки, что то делая. Из середины толпы послышался страшный крик. Петя подскакал к этой толпе, и первое, что он увидал, было бледное, с трясущейся нижней челюстью лицо француза, державшегося за древко направленной на него пики.
– Ура!.. Ребята… наши… – прокричал Петя и, дав поводья разгорячившейся лошади, поскакал вперед по улице.
Впереди слышны были выстрелы. Казаки, гусары и русские оборванные пленные, бежавшие с обеих сторон дороги, все громко и нескладно кричали что то. Молодцеватый, без шапки, с красным нахмуренным лицом, француз в синей шинели отбивался штыком от гусаров. Когда Петя подскакал, француз уже упал. Опять опоздал, мелькнуло в голове Пети, и он поскакал туда, откуда слышались частые выстрелы. Выстрелы раздавались на дворе того барского дома, на котором он был вчера ночью с Долоховым. Французы засели там за плетнем в густом, заросшем кустами саду и стреляли по казакам, столпившимся у ворот. Подъезжая к воротам, Петя в пороховом дыму увидал Долохова с бледным, зеленоватым лицом, кричавшего что то людям. «В объезд! Пехоту подождать!» – кричал он, в то время как Петя подъехал к нему.
– Подождать?.. Ураааа!.. – закричал Петя и, не медля ни одной минуты, поскакал к тому месту, откуда слышались выстрелы и где гуще был пороховой дым. Послышался залп, провизжали пустые и во что то шлепнувшие пули. Казаки и Долохов вскакали вслед за Петей в ворота дома. Французы в колеблющемся густом дыме одни бросали оружие и выбегали из кустов навстречу казакам, другие бежали под гору к пруду. Петя скакал на своей лошади вдоль по барскому двору и, вместо того чтобы держать поводья, странно и быстро махал обеими руками и все дальше и дальше сбивался с седла на одну сторону. Лошадь, набежав на тлевший в утреннем свето костер, уперлась, и Петя тяжело упал на мокрую землю. Казаки видели, как быстро задергались его руки и ноги, несмотря на то, что голова его не шевелилась. Пуля пробила ему голову.
wiki-org.ru