|

1_1 Строение клеточной мембраны | Кинезиолог
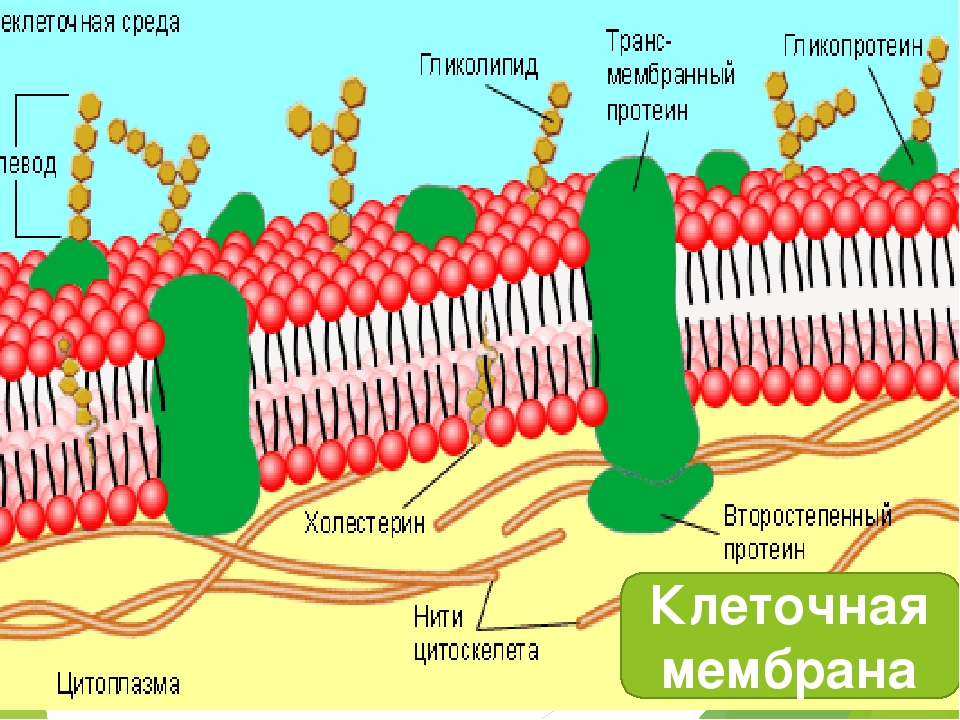
Клеточная мембрана (плазм
алемма или плазмолемма)Определение понятия
Клеточная мембрана (синонимы: плазмалемма, плазмолемма, цитоплазматическая мембрана, биомембрана) — это тройная липопротеиновая (т.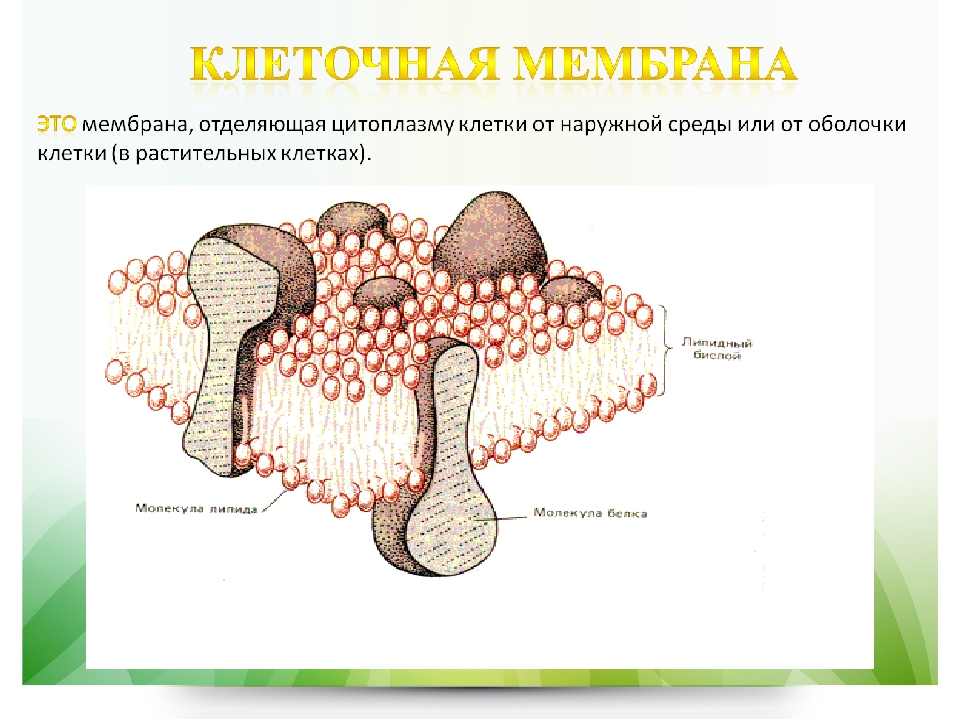 е. «жиро-белковая») оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен и связь между клеткой и окружающей её средой.
е. «жиро-белковая») оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен и связь между клеткой и окружающей её средой.
Главное в этом определении — не то, что мембрана отделяет клетку от среды, а как раз то, что она соединяет клетку с окружающей средой. Мембрана — это активная структура клетки, она постоянно работает.
Биологическая мембрана — это ультратонкая бимолекулярная пленка фосфолипидов, инкрустированная белками и полисахаридами. Эта клеточная структура лежит в основе барьерных, механических и матричных свойств живого организма (Антонов В.Ф., 1996).
Образное представление о мембране
Мне клеточная мембрана представляетсся в виде решетчатого забора с множеством дверей в нём, который окружает некую территорию. Всякая мелкая живность может через этот забор свободно перемещаться туда и обратно. Но более крупные посетители могут входить только через двери, да и то не всякие.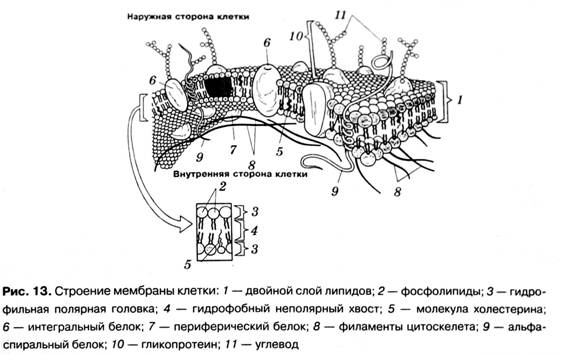 У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей — транспортных механизмов мембраны!
У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей — транспортных механизмов мембраны!
Свойства мембраны
1. Проницаемость.
2. Полупроницаемость (частичная проницаемость).
3. Избирательная (синоним: селективная) проницаемость.
4. Активная проницаемость (синоним: активный транспорт).
5. Управляемая проницаемость.
Как видим, основное свойство мембраны — это её проницаемость по отношению к различным веществам.
6. Фагоцитоц и пиноцитоз.
7. Экзоцитоз.
8. Наличие электрических и химических потенциалов, точнее разности потенциалов между внутренней и наружной сторонами мембраны. Образно можно сказать, что «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками». Подробности: смотреть тут.
Образно можно сказать, что «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками». Подробности: смотреть тут.
9. Изменения электрического и химического потенциала.
10. Раздражимость. Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лигандов (управляющих веществ). Важно отметить, что сигнальное вещество воздействует на рецептор снаружи, а изменения продолжаются внутри клетки. Получается, что мембрана передала информацию из окружающей среды во внутреннюю среду клетки.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
12. Изменение формы поверхности и её площади. Это позволяет мембране образовывать выросты наружу или, наоборот, впячивания внутрь клетки.
Это позволяет мембране образовывать выросты наружу или, наоборот, впячивания внутрь клетки.
13. Способность образовывать контакты с другими клеточными мембранами.
14. Адгезия — способность прилипать к твёрдым поверхностям.
Краткий список свойств мембраны
- Проницаемость.
- Эндоцитоз, экзоцитоз, трансцитоз.
- Потенциалы.
- Раздражимость.
- Ферментная активность.
- Контакты.
- Адгезия.
Функции мембраны
1. Неполная изоляция внутреннего содержимого от внешней среды.
2. Главное в работе клеточной мембраны — это обмен различными веществами между клеткой и межклеточной средой. Этому служит такое свойство мембраны как проницаемость. Кроме того, мембрана регулирует этот обмен за счёт того, что регулирует свою проницаемость.
3. Ещё одна важная функция мембраны — создание разности химических и электрических потенциалов между её внутренней и наружной сторонами. За счёт этого внутри клетка имеет отрицательный электрический потенциал — потенциал покоя.
За счёт этого внутри клетка имеет отрицательный электрический потенциал — потенциал покоя.
4. Через мембрану осуществляется также информационный обмен между клеткой и окружающей её средой. Специальные молекулярные рецепторы, расположенные на мембране, могут связываться с управляющими веществами (гормонами, медиаторами, модуляторами) и запускать в клетке биохимические реакции, приводящие к различным изменениям в работе клетки или в её структурах.
Видео: Строение мембраны клетки
Видеолекция: Подробно о строении мембраны и транспорте
Строение мембраны
Клеточная мембрана имеет универсальное трёхслойное строение. Её срединный жировой слой является сплошным, а верхний и нижний белковые слои покрывают его в виде мозаики из отдельных белковых участков.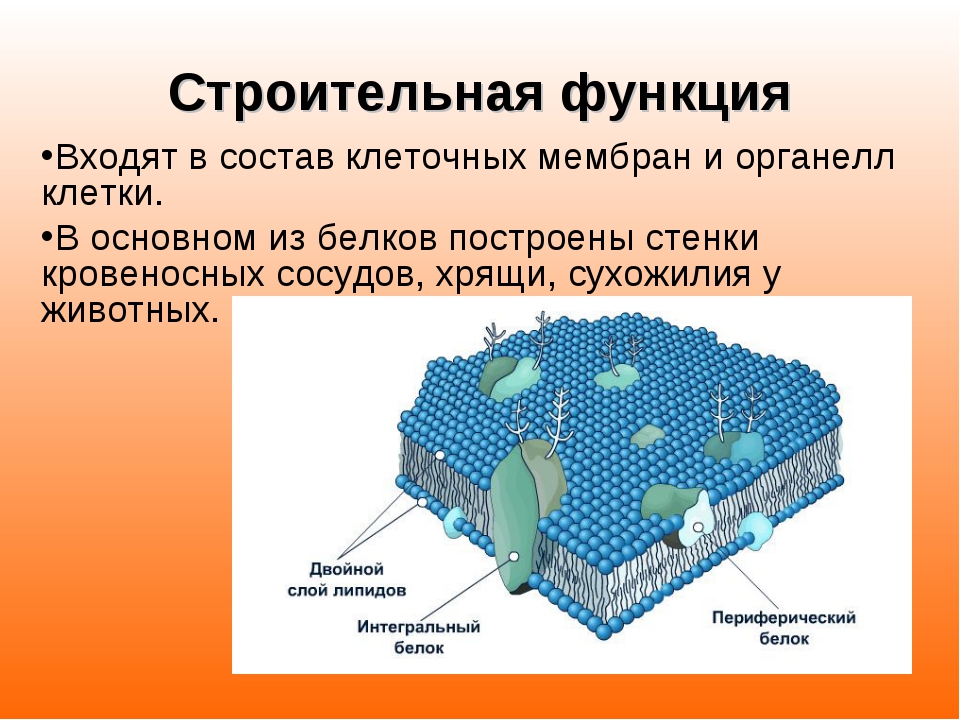 Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами — транспортёрами и ионными каналами. Зато важнейшие для всего живого газы — кислород и углекислый газ — легко перемещаются через мембрану как внутрь клетки, так и наружу.
Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами — транспортёрами и ионными каналами. Зато важнейшие для всего живого газы — кислород и углекислый газ — легко перемещаются через мембрану как внутрь клетки, так и наружу.
Ниже представлены микрофотографии реальных клеточных мембран контактирующих клеток, полученные с помощью электронного микроскопа, а также схематический рисунок, показывающий трёхслойность мембраны и мозаичность её белковых слоёв. Для увеличения изображения кликните на него.
Отдельное изображение внутреннего липидного (жирового) слоя клеточной мембраны, пронизанного интегральными встроенными белками. Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя
Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя
Рисунок выше: Неполное схематичное изображение клеточной мембраны (клеточной оболочки), приведённое в Википедии.
Учтите, что наружный и внутренний слои поверхностных белков здесь с мембраны сняты, чтобы нам лучше был виден центральный жировой двойной липидный слой. В реальной клеточной мембране сверху и снизу по жировой плёночке (мелкие шарики на рисунке) плавают большие белковые «острова», и мембрана получается более толстой, трёхслойной: белок-жир-белок. Так что она на самом деле похожа на сэндвич из двух белковых «кусков хлеба» с жирным слоем «масла» посередине, т.е. имеет трёхслойное строение, а не двухслойное.
На этом рисунке маленькие голубые и белые шарики соответствуют гидрофильным (смачиваемым) «головкам» липидов, а присоединённые к ним «ниточки» — гидрофобным (несмачиваемым) «хвостам». Из белков показаны только интегральные сквозные мембранные белки (красные глобулы и желтые спирали).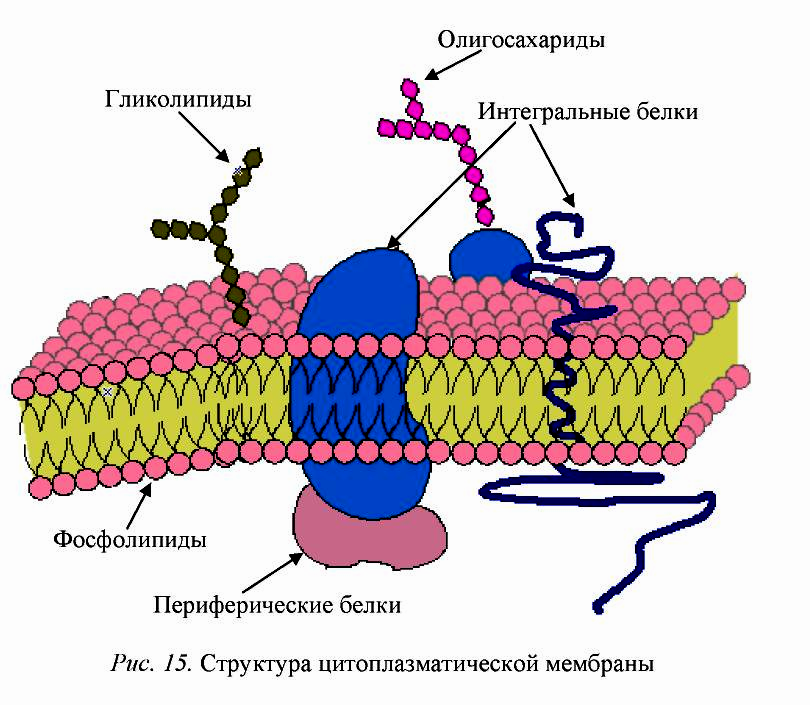 Желтые овальные точки внутри мембраны — это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс. Гликокаликс — это как бы углеводный («сахарный») «пушок» на мембране, образованный торчащими из неё длинными белково-углеводными молекулами.
Желтые овальные точки внутри мембраны — это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс. Гликокаликс — это как бы углеводный («сахарный») «пушок» на мембране, образованный торчащими из неё длинными белково-углеводными молекулами.
Модель цитоплазматической мембраны: Перейти для просмотра
Живая клетка — это маленький «белково-жировой мешочек», заполненный полужидким желеобразным содержимым, которое пронизано плёнками и трубочками.
Стенки этого мешочка образованы двойной жировой (липидной) плёночкой, облепленной изнутри и снаружи белками — клеточной мембраной. Поэтому говорят, что мембрана имеет трёхслойное строение: белки-жиры-белки. Внутри клетки также есть множество подобных жировых мембран, которые делят её внутреннее пространство на отсеки (=компартменты). Такими же мембранами окружены клеточные органеллы: ядро, митохондрии, хлоропласты. Так что мембрана — это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.
Так что мембрана — это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.
Слева — уже не реальная, а искусственная модель кусочка биологической мембраны: это мгновенный снимок жирового фосфолипидного бислоя (т.е. двойного слоя) в процессе его молекулярно-динамического моделирования. Показана расчётная ячейка модели — 96 молекул ФХ (фосфатидилхолина) и 2304 молекулы воды, всего 20544 атомов.
Справа — наглядная модель одиночной молекулы того самого липида, из которых как раз и собирается мембранный липидный бислой. Вверху у него гидрофильная (водолюбивая) головка, а снизу — два гидрофобных (боящихся воды) хвостика. У этого липида есть простое название: 1-стероил-2-докозагексаеноил-Sn-глицеро-3-фосфатидилхолин (18:0/22:6(n-3)cis ФХ), но вам нет нужды его запоминать, если вы только не планируете довести своего преподавателя до обморока глубиной своих познаний.
Можно дать и более точное научное определение клетке:
Клетка – это ограниченная активной мембраной, упорядоченная, структурированная неоднородная система биополимеров, участвующих в единой совокупности обменных, энергетических и информационных процессов, и также осуществляющих поддержание и воспроизведение всей системы в целом.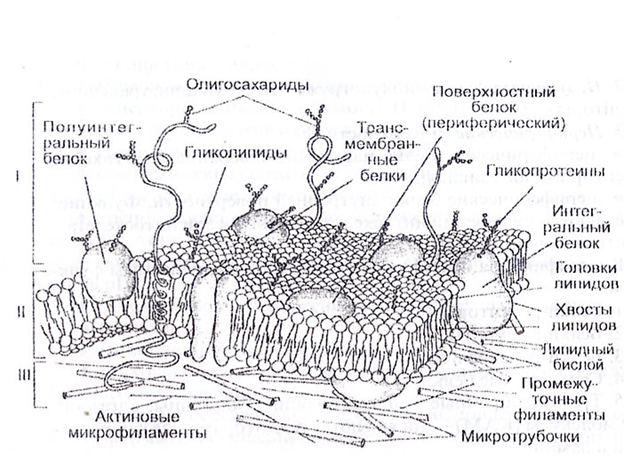
Внутри клетка также пронизана мембранами, а между мембранами находится не вода, а вязкий гель/золь изменяемой плотности. Поэтому взаимодействующие молекулы в клетке не плавают свободно, как в пробирке с водным раствором, а в основном сидят (иммобилизованы) на полимерных структурах цитоскелета или внутриклеточных мембранах. И химические реакции поэтому проходят внутри клетки почти как в твердом теле, а не в жидкости. Наружная мембрана, окружающая клетку, также облеплена ферментами и молекулярными рецепторами, что делает её очень активной частью клетки.
Клеточная мембрана (плазмалемма, плазмолемма) — это активная оболочка, отделяющая клетку от окружающей среды и связывающая её с окружающей средой. © Сазонов В.Ф., 2016.
Из этого определения мембраны следует, что она не просто ограничивает клетку, а активно работает, связывая её с окружающей её средой.
Мембранные липиды
Жир, из которого состоят мембраны, — особенный, поэтому его молекулы принято называть не просто жиром, а «липидами», «фосфолипидами», «сфинголипидами».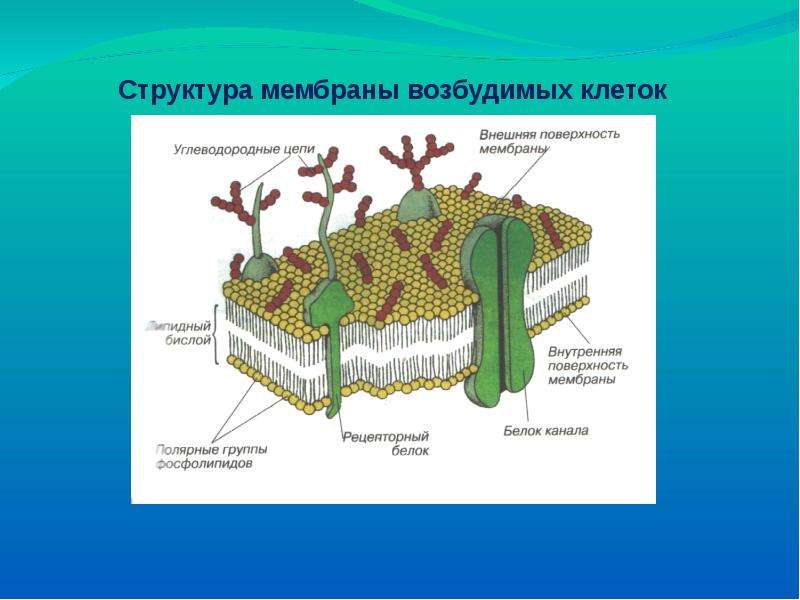
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин, а также в меньших количествах гликолипиды.
С химической точки зрения фосфолипид состоит из четырёх частей: глицерина, двух жирных кислот с длинной углеводородной цепью, фосфорной кислоты и особой для каждого фосфолипида группы, которую принято называть характеристической группой. Трёхатомный спирт глицерин связывает через сложно-эфирную связь две жирные кислоты и остаток фосфорной кислоты, к которой присоединена характеристическая группа (например, этаноламин).
Рис. ___. Структурная формула фосфатидилэтаноламина как пример амфифильной (гидрофобной/гидрофильной) молекулы фосфолипида. Кроме этаноламина характеристической группой фосфолипида может быть также холин, инозитол, серин и некоторые другие молекулы.
Рис. ___. Молекулярная структура фосфатидилхолина (=лецитина). Источник изображения: https://pandia.ru/text/80/650/73429-4.php
Мембранная плёночка является двойной, т. е. она состоит из двух липидных плёночек, слипшихся друг с другом с помощью своих липидных «хвостиков». Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из «бислоя«, т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая — не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами. Примерно так можно соединить две щётки, направив их щетиной друг к другу и слегка придавив.
е. она состоит из двух липидных плёночек, слипшихся друг с другом с помощью своих липидных «хвостиков». Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из «бислоя«, т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая — не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами. Примерно так можно соединить две щётки, направив их щетиной друг к другу и слегка придавив.
Мембранные белки
Белки мембраны включены в липидный двойной слой двумя способами:
- Гидрофильные радикалы аминокислот поверхностных мембранных белков связаны нековалентными связями с гидрофильной поверхностью липидного бислоя.
- Интегральные мембранные белки погружены в гидрофобную область бислоя.
Интегральные белки различаются по степени погруженности в гидрофобную часть бислоя. Они могут располагаться по обеим сторонам мембраны и при этом либо частично погружаются в мембрану, либо располагаются трансмембранно.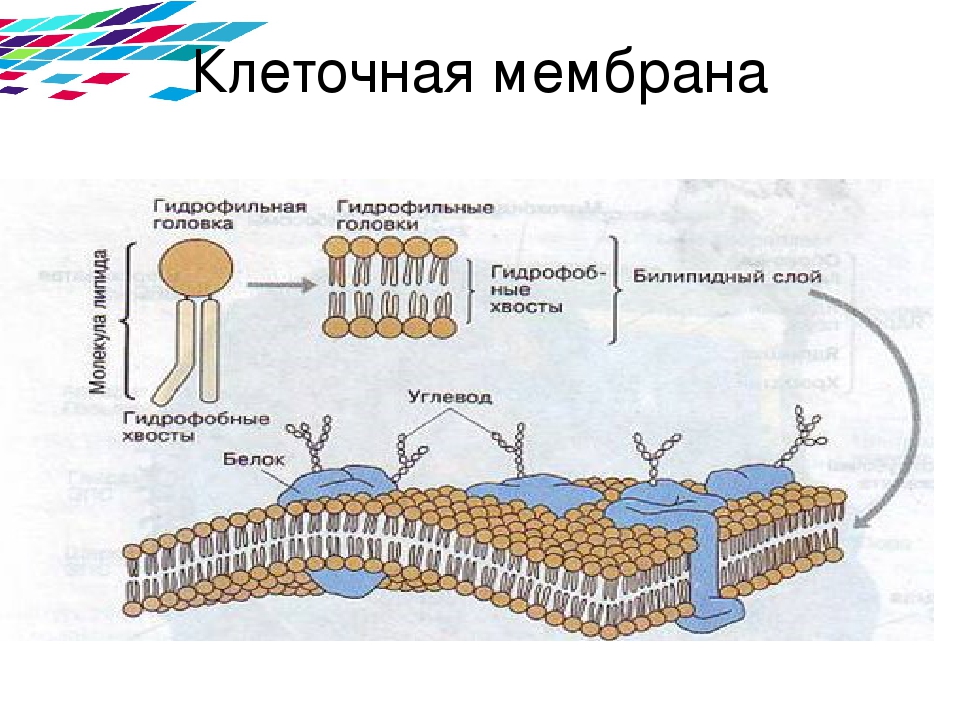 Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связана с моносахаридными остатками или олигосахаридными цепями и представляет собой гликопротеины. В отличие от нерастворимых фибриллярных белков растворимые белки имеют почти сферическую (глобулярную) форму. Глобулярным белкам свойственна высокоупорядоченная пространственная структура (конформация), которая способствует выполнению специфических биологических функций (Албертс и соавт., 1994).
Погруженная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связана с моносахаридными остатками или олигосахаридными цепями и представляет собой гликопротеины. В отличие от нерастворимых фибриллярных белков растворимые белки имеют почти сферическую (глобулярную) форму. Глобулярным белкам свойственна высокоупорядоченная пространственная структура (конформация), которая способствует выполнению специфических биологических функций (Албертс и соавт., 1994).
Подвижными в мембране являются не только липиды, но и мембранные белки. Если белки не закреплены в мембране, они «плавают» в липидном бислое как в жидкости. Поэтому говорят, что биомембраны имеют жидкостно-мозаичную структуру.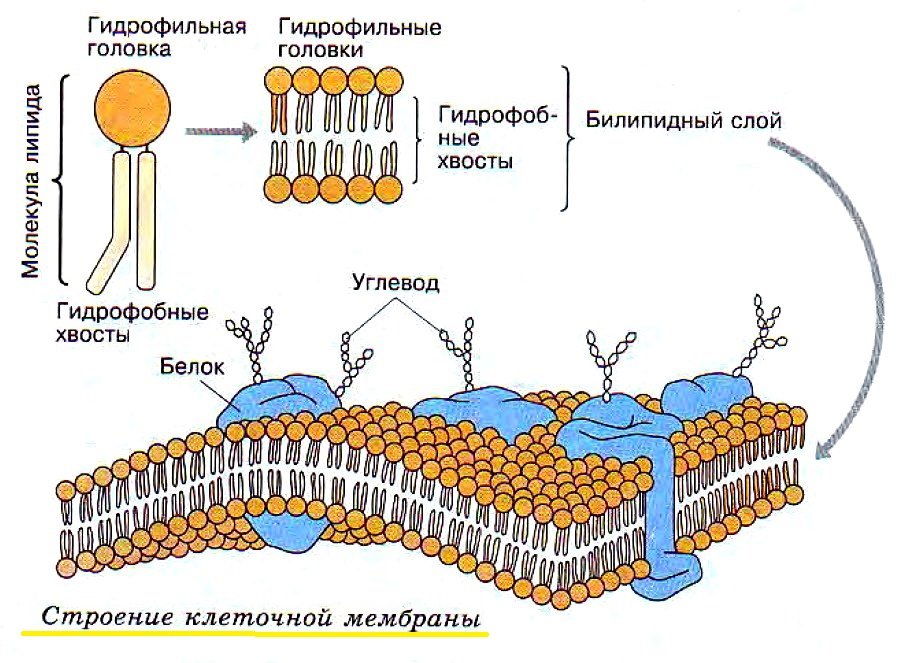 При этом «дрейф» белков в плоскости мембраны происходит достаточно легко, переход их с внешней стороны мембраны на внутреннюю («флип-флоп») невозможен, а переход липидов происходит крайне редко. Для «перескока» липидов необходимы специальные белки транслокаторы. Исключение составляет жир холестерин, который может легко переходить с одной стороны мембраны на другую. Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением. Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря» и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками. У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21–25 преимущественно гидрофобных аминокислот, которые образуют правую трансмембранную α-спираль с 6 или 7 витками (Фалер, Шилдс, 2004).
При этом «дрейф» белков в плоскости мембраны происходит достаточно легко, переход их с внешней стороны мембраны на внутреннюю («флип-флоп») невозможен, а переход липидов происходит крайне редко. Для «перескока» липидов необходимы специальные белки транслокаторы. Исключение составляет жир холестерин, который может легко переходить с одной стороны мембраны на другую. Интегральные мембранные белки имеют трансмембранные спирализованные участки (домены), которые однократно или многократно пересекают липидный бислой. Такие белки прочно связаны с липидным окружением. Периферические мембранные белки удерживаются на мембране с помощью липидного «якоря» и связаны с другими компонентами мембраны; например, они часто бывают ассоциированы с интегральными мембранными белками. У интегральных мембранных белков фрагмент пептидной цепи, пересекающий липидный бислой, обычно состоит из 21–25 преимущественно гидрофобных аминокислот, которые образуют правую трансмембранную α-спираль с 6 или 7 витками (Фалер, Шилдс, 2004).
Мембрана бактерий
Оболочка прокариотической клетки грамотрицательных бактерий состоит из нескольких слоёв, показанных на рисунке ниже.
Слои оболочки грамотрицательных бактерий:
1. Внутренняя трёхслойная цитоплазматическая мембрана, которая соприкасается с цитоплазмой.
2. Клеточная стенка, которая состоит из муреина.
3. Наружная трёхслойная цитоплазматическая мембрана, которая имеет такую же систему липидов с белковыми комплексами, как и внутренняя мембрана.
Общение грамотрицательных бактериальных клеток с внешним миром через такую сложную трёхступенчатую структуру не даёт им преимущества в выживании в суровых условиях по сравнению с грамположительным бактериями, имеющими менее мощную оболочку. Они точно так же плохо переносят высокие температуры, повышенную кислотность и перепады давления.
Рис. Сложная тройная клеточная оболочка грамотрицательных бактерий. Источник изображения: https://probakterii.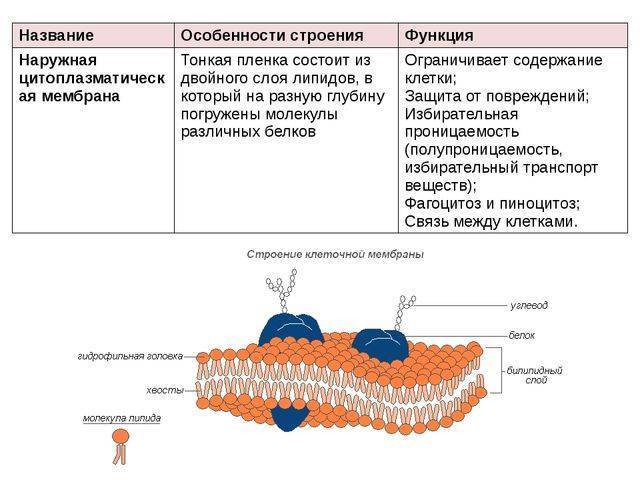 ru/prokaryotes/organelles/membrana-bakterij.html
ru/prokaryotes/organelles/membrana-bakterij.html
Рис. Сравнение оболочек грамположительных и грамотрицательных бактерий. Источник изображения: https://myslide.ru/presentation/512325_skachat-stroenie-bakterialnoj-kletki
Рис . Рафтовые неоднородности в мембране различного масштаба. а — Нанокластеры холестерола, сфингомиелина, гликосфинголипидов и белков плазматической мембраны различаются по составу. Считается, что в эти кластеры входят ГФИ-заякоренные белки, трансмембранные (ТМ) белки, специфичные для рафтов, и цитоплазматические белки, связанные с актиновыми филаментами. «Обычные» ТМ-белки не входят в состав рафтов. б — В ответ на внешние сигналы нанокластеры могут сливаться с образованием рафтовой платформы, важной для ТМ передачи сигналов и мембранного транспорта. в — Рафтовая фаза, видимая в микроскоп (ø ≈1 мкм), наблюдается исключительно в равновесных мембранных системах, таких как гигантские синтетические или мембранные везикулы.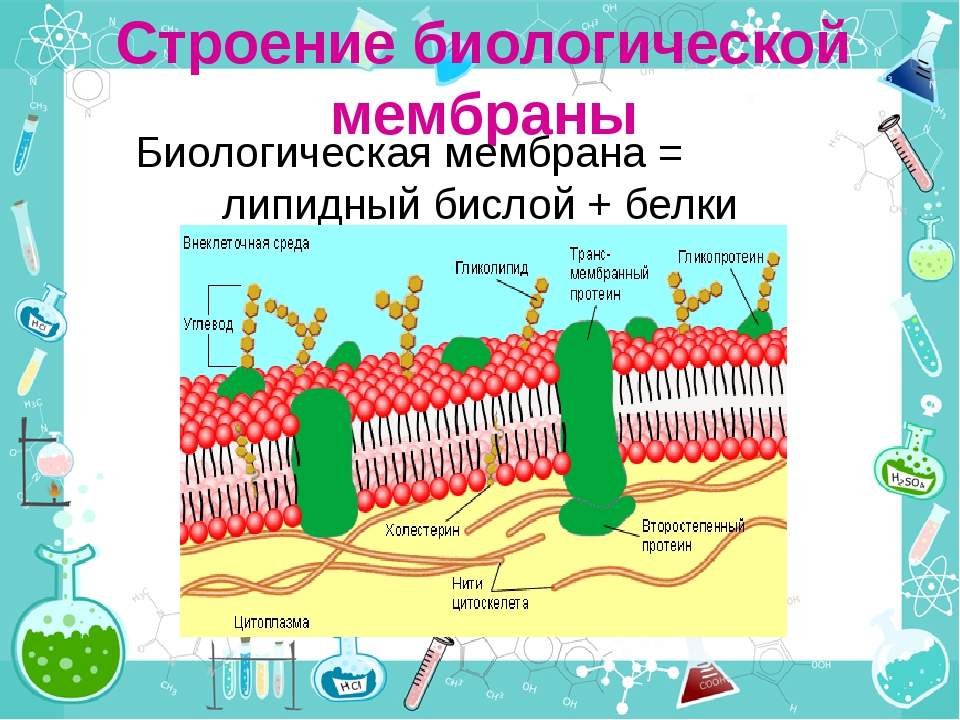 В «нативных» мембранах постоянный обмен веществом и энергией «дробит» рафтовую фазу до субдифракционных размеров…. Читайте дальше на Биомолекуле: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni Источник изображения: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni
В «нативных» мембранах постоянный обмен веществом и энергией «дробит» рафтовую фазу до субдифракционных размеров…. Читайте дальше на Биомолекуле: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni Источник изображения: https://biomolecula.ru/articles/lipidnyi-fundament-zhizni
Рис. Domain-length scales and the biomembrane as a protein–lipid composite material. (a) Length scales of domains in biomembranes. Shells, complexes and nanoclusters range from 1–10 nm, whereas nanodomains such as caveolae can be as large as 100 nm. (b) A schematic representation of the biomembrane as a composite of lipids and proteins. Estimates of lateral protein concentration are about 30,000 per μm2 based on rhodopsin in the rod outer segment28,29 and transmembrane proteins in the baby hamster kidney (BHK) cell membrane27. Lipids were assumed to occupy a surface area of ∼0.68 nm2 (diameter ∼0.93 nm) and an α-helix ∼1 nm2 (diameter ∼1.1 nm). A 30 × 30 nm2 section of membrane is depicted with 32 lipids on a side, 35 transmembrane proteins with 15 single-span, 12 tetraspan and eight heptaspan α-helical proteins, having assumed crosssectional areas in the plane of the membrane of 1 nm2, 4. 5 nm2 and 8 nm2, respectively. Taking into account the area excluded by the proteins, the numerical lipid : protein ratio is ∼50. For a single-span helix with a diameter of ∼1.1 nm, there are about seven lipids in the first boundary layer; for a tetraspan protein with a diameter of ∼2.4 nm, there are about 11 lipids in the first boundary layer; for a heptaspan protein (such as rhodopsin) with a diameter of ∼3.2 nm, there would be about 14 lipids in the first boundary layer. Such first-boundary layer lipids are shown in white, whereas the second layer is shown in red. All other lipids are shown in yellow. Lipid-binding proteins and adaptors linking transmembrane proteins to membrane proximate cytoskeletal filaments are also depicted as different coloured structures beneath the plane of the membrane, but ectodomains of the membrane proteins are omitted for clarity. Источник изображения: https://www.nature.com/articles/ncb0107-7
5 nm2 and 8 nm2, respectively. Taking into account the area excluded by the proteins, the numerical lipid : protein ratio is ∼50. For a single-span helix with a diameter of ∼1.1 nm, there are about seven lipids in the first boundary layer; for a tetraspan protein with a diameter of ∼2.4 nm, there are about 11 lipids in the first boundary layer; for a heptaspan protein (such as rhodopsin) with a diameter of ∼3.2 nm, there would be about 14 lipids in the first boundary layer. Such first-boundary layer lipids are shown in white, whereas the second layer is shown in red. All other lipids are shown in yellow. Lipid-binding proteins and adaptors linking transmembrane proteins to membrane proximate cytoskeletal filaments are also depicted as different coloured structures beneath the plane of the membrane, but ectodomains of the membrane proteins are omitted for clarity. Источник изображения: https://www.nature.com/articles/ncb0107-7
Видеолекция: Плазматическая мембрана. Е.В. Шеваль, к.б.н.
Е.В. Шеваль, к.б.н.
Видеолекция: Мембрана как клеточная граница. А. Иляскин
Важность ионных каналов мембраны
Легко понять, что через мембранную жировую плёнку могут проникать в клетку только жирорастворимые вещества. Это жиры, спирты, газы. Например, в эритроцитх прямо через мембрану легко проходят внутрь и наружу кислород и углекислый газ. А вот вода и водорастворимые вещества (например, ионы) просто так через мембрану не могут пройти внутрь любой клетки. Это значит, что для них нужны специальные отверстия. Но если просто сделать отверстие в жировой плёнке, то оно тут же затянется обратно. Что же делать? Выход в природе был найден: надо сделать специальные белковые транспортные структуры и протянуть их сквозь мембрану. Именно так и получаются каналы для пропускания не растворимых в жире веществ — ионные каналы мембраны клетки.
Именно так и получаются каналы для пропускания не растворимых в жире веществ — ионные каналы мембраны клетки.
Итак, для придания своей мембране дополнительных свойства проницаемости для полярных молекул (ионов и воды) клетка синтезирует в цитоплазме специальные белки, которые затем встраиваются в мембрану. Они бывают двух типов: белки-транспортёры (например, транспортные АТФазы) и белки-каналоформеры (образователи каналов). Эти белки встраиваются в двойной жировой слой мембраны и формируют транспортные структуры в виде транспортёров или в виде ионных каналов. Через эти транспортные структуры теперь могут проходить различные водорастворимые вещества, которые по-другому проходить сквозь жировую мембранную плёнку не могут.
Вообще, встроенные в мембрану белки ещё называются интегральными, именно потому что они как бы включаются в состав мембраны и пронизывают её насквозь. Другие белки, не интегральные, образуют как бы острова, «плавающие» по поверхности мембраны: либо по её наружной поверхности, либо по внутренней. Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко!
Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко!
Выводы
1. В целом, мембрана получается трёхслойной:
1) наружный слой из белковых «островов»,
2) жировое двухслойное «море» (липидный бислой), т.е. двойная липидная плёнка,
3) внутренний слой из белковых «островов».
Но есть ещё рыхлый наружный слой — гликокаликс, который образуют торчащие из мембраны гликопротеины. Они являются молекулярными рецепторами, с которыми связываются сигнальные управляющие вещества.
2. В мембрану встроены специальные белковые структуры, обеспечивающие её протицаемость для ионов или других веществ. Не надо забывать, что в некоторых местах жировое море пронизано интегральными белками насквозь. И именно интегральные белки образуют специальные транспортные структуры клеточной мембраны (смотрите раздел ). Через них вещества попадают внутрь клетки, а также выводятся из клетки наружу.
3. С любой стороны мембраны (наружной и внутренней), а также внутри мембраны могут располагаться белки-ферменты, которые влияют и на состояние самой мембраны и на жизнь всей клетки.
С любой стороны мембраны (наружной и внутренней), а также внутри мембраны могут располагаться белки-ферменты, которые влияют и на состояние самой мембраны и на жизнь всей клетки.
Так что мембрана клетки — это активная изменчивая структура, которая активно работает в интересах всей клетки и связывает её с окружающим миром, а не просто является «защитной оболочкой». Это — самое важное, что надо знать про клеточную мембрану.
В медицине мембранные белки зачастую используются как “мишени” для лекарственных средств. В качестве таких мишеней выступают рецепторы, ионные каналы, ферменты, транспортные системы. В последнее время кроме мембраны мишенью для лекарственных веществ становятся также гены, спрятанные в клеточном ядре.
Видео: Введение в биофизику клеточной мембраны: Структура мембран 1 (Владимиров Ю.А.)
Видео: История, строение и функции клеточной мембраны: Структура мембран 2 (Владимиров Ю. А.)
А.)
Дополнительно: Антонов В.Ф., 1996.
Подробности о биомембранах на сайте Биомолекула
Читать далее:
© 2010-2021 Сазонов В.Ф. © 2010-2016 kineziolog.bodhy.ru, © 2016-2021 kineziolog.su
Клеточная мембрана
☰
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO 2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами).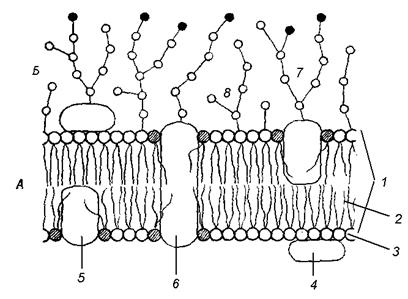 Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.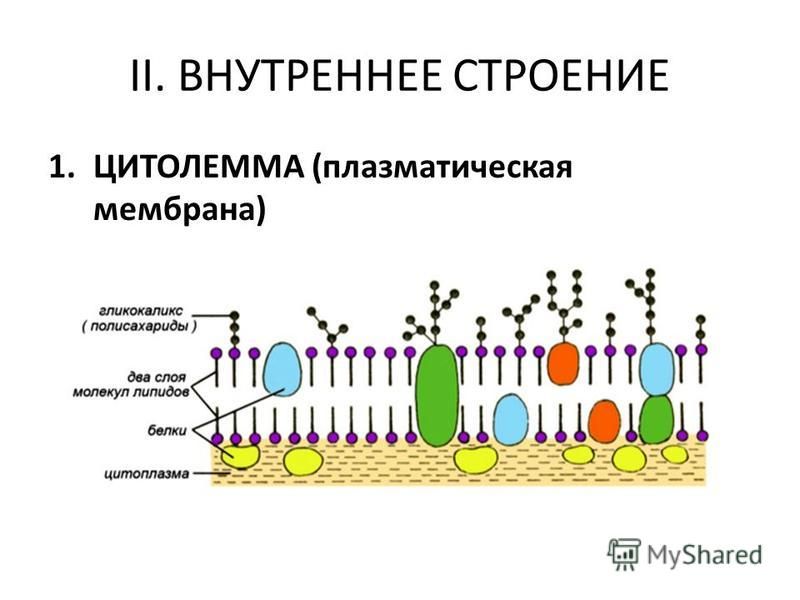
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Строение клеточной мембраны
☰
Клеточная мембрана имеет достаточно сложное строение, которое можно рассмотреть в электронный микроскоп. Грубо говоря, она состоит из двойного слоя липидов (жиров), в который в разных местах включены различные пептиды (белки). Общая толщина мембраны составляет около 5-10 нм.
Общий план строения клеточной мембраны универсален для всего живого мира. Однако мембраны животных содержат включения холестерина, который определяет ее жесткость.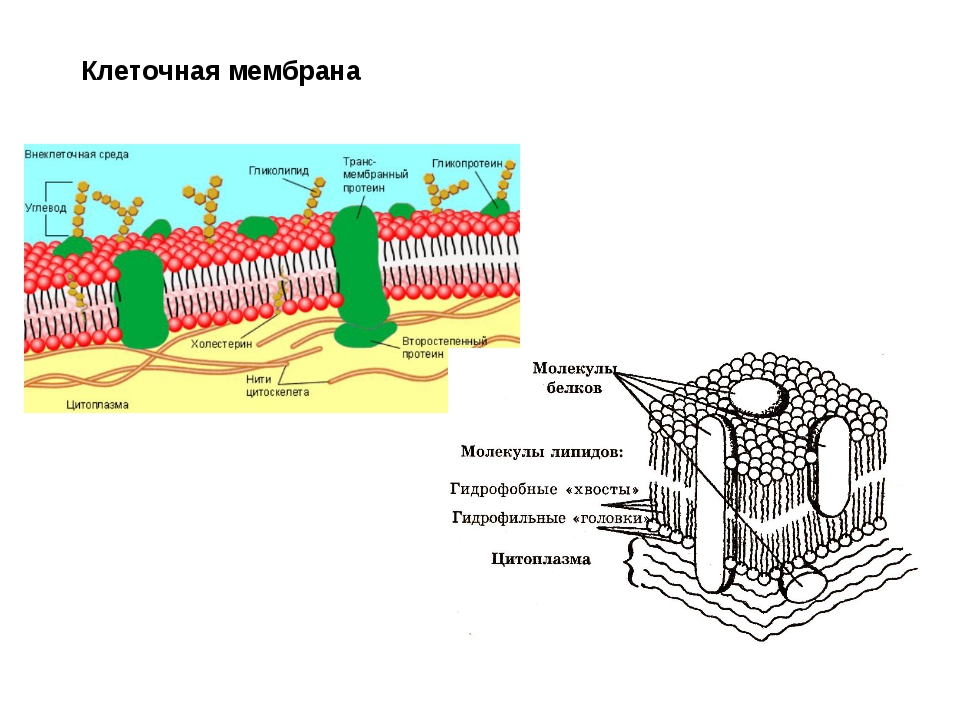 Отличие мембран разных царств организмов в основном касается надмембранных образований (слоев). Так у растений и грибов над мембраной (с внешней стороны) находится клеточная стенка. У растений она состоит преимущественно из целлюлозы, а у грибов — из вещества хитина. У животных надмембранный слой называется гликокаликсом.
Отличие мембран разных царств организмов в основном касается надмембранных образований (слоев). Так у растений и грибов над мембраной (с внешней стороны) находится клеточная стенка. У растений она состоит преимущественно из целлюлозы, а у грибов — из вещества хитина. У животных надмембранный слой называется гликокаликсом.
По-другому клеточная мембрана называется цитоплазматической мембраной или плазматической мембраной.
Более глубокое изучение строения клеточной мембраны открывает многие ее особенности, связанные с выполняемыми функциями.
Двойной слой липидов в основном состоит из фосфолипидов. Это жиры, один конец которых содержит остаток фосфорной кислоты, обладающий гидрофильными свойствами (т. е. притягивает молекулы воды). Второй конец фосфолипида — это цепи жирных кислот, обладающие гидрофобными свойствами (не образуют с водой водородных связей).
Молекулы фосфолипидов в клеточной мембране выстраиваются в два ряда так, что их гидрофобные «концы» находятся внутри, а гидрофильные «головки» – снаружи.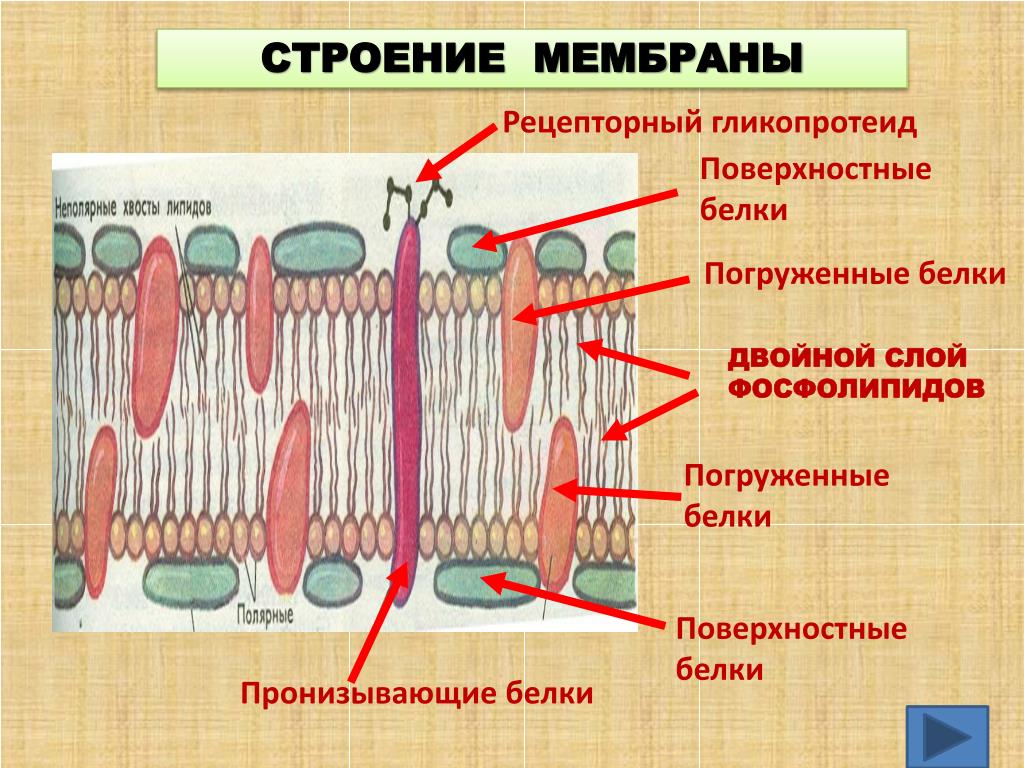 Получается достаточно прочная структура, ограждающая содержимое клетки от внешней среды.
Получается достаточно прочная структура, ограждающая содержимое клетки от внешней среды.
Белковые включения в клеточной мембране распределены неравномерно, кроме того они подвижны (так как фосфолипиды в бислое обладают боковой подвижностью). С 70-х годов XX века стали говорить о жидкостно-мозаичном строении клеточной мембраны.
В зависимости от того, как белок входит в состав мембраны, выделяют три типа белков: интегральные, полуинтегральные и периферические. Интегральные белки проходят через всю толщу мембраны, и их концы торчат по обеим ее сторонам. В основном выполняют транспортную функцию. У полуинтегральных белков один конец находится в толще мембраны, а второй выходит наружу (с внешней или внутренней) стороны. Выполняют ферментативную и рецепторную функции. Периферические белки находятся на внешней или внутренней поверхности мембраны.
Особенности строения клеточной мембраны говорят о том, что она является основным компонентом поверхностного комплекса клетки, но не единственным. Другими его компонентами являются надмембранный слой и субмембранный слой.
Другими его компонентами являются надмембранный слой и субмембранный слой.
Гликокаликс (надмембранный слой животных) образуют олигосахариды и полисахариды, а также периферические белки и выступающие части интегральных белков. Компоненты гликокаликса выполняют рецепторную функцию.
Кроме гликокаликса у клеток животных бывают и другие надмембранные образования: слизи, хитин, перилемма (подобна мембране).
Надмембранным образованием у растений и грибов является клеточная стенка.
Субмембранный слой клетки — это поверхностная цитоплазма (гиалоплазма) с входящей в нее опорно-сократительной системой клетки, фибриллы которой взаимодействуют с белками, входящими в клеточную мембрану. По таким соединениям молекул передаются различные сигналы.
Структура, функции и роль клеточной мембраны
Автор Nat WorldВремя чтения 3 мин.Просмотры 683Опубликовано Обновлено
Клеточная мембрана (плазматическая мембрана) представляет собой тонкую полупроницаемую оболочку, которая окружает цитоплазму клетки.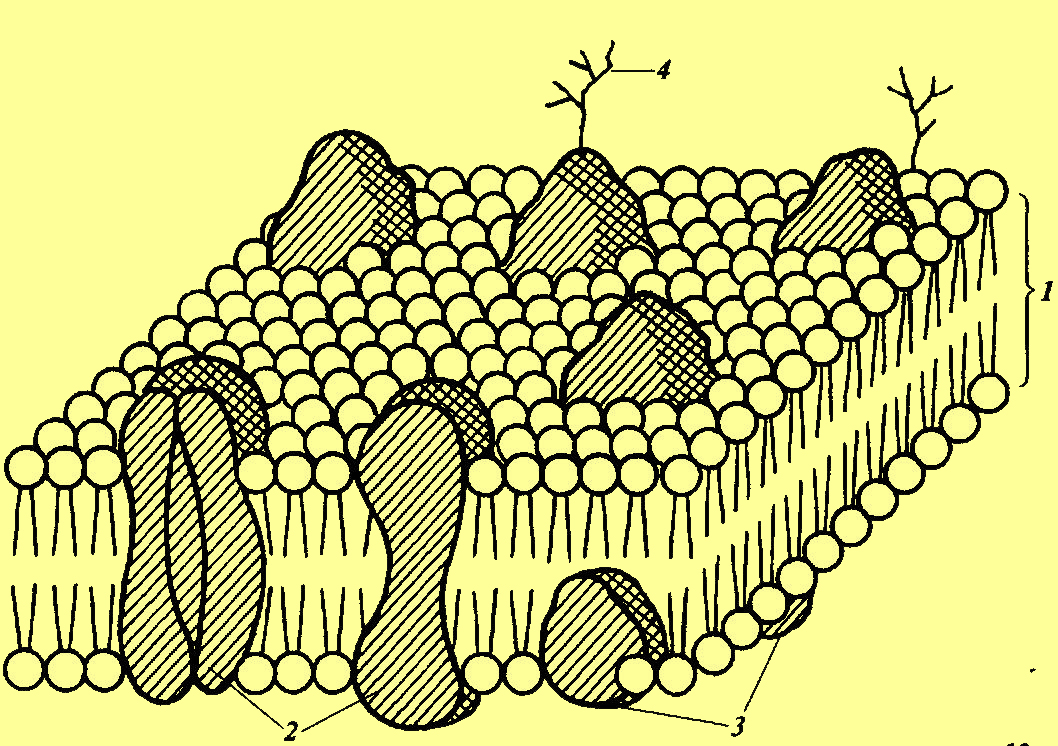
Функция и роль клеточной мембраны
Ее функция заключается в том, чтобы защитить целостность внутренней части клетки, впуская некоторые необходимые вещества в клетку, и не позволяя проникать другим.
Он также служит основой привязанности к цитоскелету у одних организмов и к клеточной стенке у других. Таким образом, плазматическая мембрана также обеспечивает форму клетки. Еще одна функция мембраны заключается в регулировании роста клеток через баланс эндоцитоза и экзоцитоза.
При эндоцитозе липиды и белки удаляются из клеточной мембраны по мере усвоения веществ. При экзоцитозе везикулы, содержащие липиды и белки, сливаются с клеточной мембраной, увеличивая размер клеток. Животные, растительные и грибковые клетки имеют плазматические мембраны. Внутренние органеллы, например, ядро, также заключены в защитные мембраны.
Структура клеточной мембраны
Плазматическая мембрана в основном состоит из смеси белков и липидов. В зависимости от расположения и роли мембраны в организме, липиды могут составлять от 20 до 80 процентов мембраны, а остальная часть приходится на белки. В то время как липиды помогают придать мембране гибкость, белки контролируют и поддерживают химический состав клетки, а также помогают в переносе молекул сквозь мембрану.
В то время как липиды помогают придать мембране гибкость, белки контролируют и поддерживают химический состав клетки, а также помогают в переносе молекул сквозь мембрану.
Липиды мембран
Фосфолипиды являются основным компонентом плазматических мембран. Они образуют липидный бислой, в котором гидрофильные (притянутые к воде) участки «головы» спонтанно организуются, чтобы противостоять водному цитозолю и внеклеточной жидкости, тогда как гидрофобные (отталкиваемые водой) участки «хвоста» обращены от цитозоля и внеклеточной жидкости. Липидный бислой является полупроницаемым, позволяя только некоторым молекулам диффундировать через мембрану.
Холестерин является еще одним липидным компонентом мембран животных клеток. Молекулы холестерина избирательно диспергированы между мембранными фосфолипидами. Это помогает сохранить жесткость клеточных мембран, предотвращая слишком плотное расположение фосфолипидов. Холестерин отсутствует в мембранах растительных клеток .
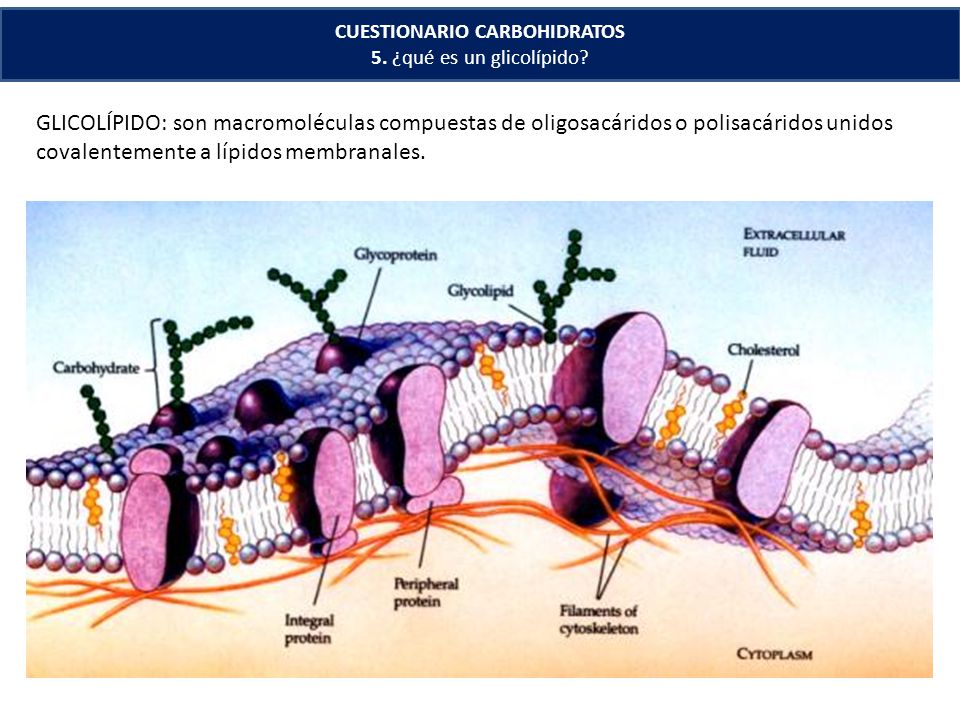
Гликолипиды расположены с наружной поверхности клеточных мембран и соединяются с ними углеводной цепью. Они помогают клетке распознавать другие клетки организма.
Они помогают клетке распознавать другие клетки организма.
Белки мембран
Клеточная мембрана содержит два типа ассоциированных белков. Белки периферической мембраны являются внешними и связаны с ней путем взаимодействия с другими белками. Интегральные мембранные белки вводятся в мембрану, и большинство проходит сквозь нее. Части этих трансмембранных белков расположены по обе ее стороны.
Белки плазматической мембраны имеют ряд различных функций. Структурные белки обеспечивают поддержку и форму клеток. Белки рецептора мембраны помогают клеткам контактировать со своей внешней средой с помощью гормонов, нейротрансмиттеров и других сигнальных молекул. Транспортные белки, такие как глобулярные белки, переносят молекулы через клеточные мембраны посредством облегченной диффузии. Гликопротеины имеют прикрепленную к ним углеводную цепь. Они встроены в клеточную мембрану, помогая в обмене и переносе молекул.
Мембраны органелл
Некоторые клеточные органеллы также окружены защитными мембранами. Ядро, эндоплазматический ретикулум, вакуоль, лизосома и аппарат Гольджи являются примерами окруженных мембраной органелл. Митохондрии и хлоропласты покрыты двойной мембраной. Мембраны различных органелл различаются по молекулярному составу и хорошо подходят для выполнения своей роли. Они важны для нескольких жизненно важных функций клеток, включая синтез белка, производство липидов и клеточное дыхание.
Ядро, эндоплазматический ретикулум, вакуоль, лизосома и аппарат Гольджи являются примерами окруженных мембраной органелл. Митохондрии и хлоропласты покрыты двойной мембраной. Мембраны различных органелл различаются по молекулярному составу и хорошо подходят для выполнения своей роли. Они важны для нескольких жизненно важных функций клеток, включая синтез белка, производство липидов и клеточное дыхание.
Не все нашли? Используйте поиск по сайту ↓
Что такое клеточная мембрана? / Справочник :: Бингоскул
Каждая клеточка многоклеточного организма отделена от окружающей внутренней (или наружной) среды плазматической мембраной (цитолеммой, плазмалеммой) Это граница, которая «охраняет» внутреннее содержимое клетки от чужеродного вторжения. Но, как и на любой границе, она пропускает «своих» и выводит « чужих». Мембрана – это не панцирь, а подвижная, активная структура, благодаря которой осуществляется обмен веществ между клетками и окружающей средой.
Рис. 1. Клеточная мембранаВ переводе с латыни слово «мембрана» означает «пленка». Клетка завернута или упакована со всех сторон оболочкой, строение которой уникально. Она подвижна: тонкая, гибкая и эластичная диаметром 7,5-10 нм.
Клеточная мембрана похожа на гамбургер, состоящий из трех слоев: липиды – белки – липиды. Но это условное сравнение. На фото видно, что устроена она более сложно: два тонких липидных слоя пронизаны белковыми глобулами, которые встроены в оболочку. Вывод: мембрана подвижна и состоит из белков и липидов (жиров). Это динамичная структура, где липиды и белки способны перемещаться, меняя свое положение.
Строение
С 1925 года, когда впервые рассмотрели мембрану, гипотеза о ее строении претерпела ряд существенных изменений. С 1972 года общепризнанной считается жидкостно-мозаичная модель плазматической мембраны, которая представлена на схеме:
Рис. 2. Жидкостно-мозаичная модель мембраныФосфолипидный биослой устроен таким образом, что гидрофобные остатки жирных кислот обращены внутрь, а гидрофильные головки «смотрят» наружу. Двойной липидный слой – это барьер, благодаря которому содержимое клетки не растекается, и во внутреннее содержимое не попадают токсические вещества.
Рис. 3. Строение клеточной мембраныВ 2-хслойный липидный слой погружены подвижные молекулы белков, которые по выполняемым функциям делятся на 2 большие группы:
- Структурные, которые в прямом смысле поддерживают структуру клетки.
- Динамические, находящиеся на поверхности мембраны. Они участвуют в «барьерных» процессах. Среди динамических различают рецепторные, транспортные и ферментативные белковые глобулы.
На заметку: Соотношение компонентов в плазматической мембране следующее: белки составляют 65%, на фосфолипиды приходится 25%, холестерол – 13%, липиды другого строения – 4%, углеводы – 3%. Липиды (жиры) отталкивают воду, поэтому ее излишки не попадают в клетку.
Функции клеточной мембраны
Плазматическая мембрана ведет себя активно, как любой клеточный органоид. Это не просто барьер: «клеточная капсула» выполняет много задач.
Транспортная функция
Рис. 4. Механизмы прохождения веществ через клеточную мембрануОсуществляет выборочный транспорт веществ, поскольку липидные слои для большинства соединений непроницаемы. Через мембрану идет пассивный и активный транспорт:
1. Пассивный транспорт — это простая диффузия, которая идет с низкой скоростью. Через барьер проникают газы, вода, ряд органических веществ. Облегченная диффузия происходит с участием белков-переносчиков и идет с большей скоростью. На пассивный транспорт не требуется энергетических затрат. Белки-переносчики относятся к транспортным. Они «проносят» вещества через мембрану или через специальные каналы, предназначенные для проникновения различных ионов.
2. Перенос макромолекул или крупных частиц осуществляется путем эндоцитоза. При эндоцитозе на мембране возникают впячивания для захвата твердой частицы пищи. Вокруг нее образуется вакуоль, которая обволакивает частицу и переносит внутрь клетки.
Рис. 5. Эндоцитоз
На заметку: Эндоцитоз делится на фагоцитоз или захват твердой частицы и пиноцитоз, когда поглощается жидкий материал (коллоидный раствор, суспензии или просто раствор).
3. Вывод веществ из клетки через мембрану осуществляется путем экзоцитоза. Вещества, которые нужно переправить в межклеточную жидкость, «упаковываются» в мембранные пузырьки. Они подходят к цитолемме, встраиваются в нее и содержимое пузырьков выбрасывается из клетки. Это различные продукты метаболизма.
Рис. 6. Экзоцитоз4. Активный транспорт осуществляется против градиента концентрации и требует затрат энергии (АТФ). Вещества из области с меньшей концентрацией переходят в область с более концентрированным содержимым. Примером служит натрий-калиевый насос, когда из клетки выводятся ионы натрия и закачиваются ионы калия.
Проницаемость мембран разных клеток неодинакова. Чтобы попасть в клетку, вещество должно обладать определенным размером, химическими свойствами и электрическим зарядом. «Неподходящие» по своим параметрам молекулы просто не способны попасть во внутреннее содержимое клетки.
Другие функции клеточной мембраны
Плазматическая мембрана осуществляет:
- Структурную функцию, отделяя клетку от внешней среды. Так клетка работает как автономный организм. Она содержит набор органоидов, которые «плавают» во внутренней среде – цитоплазме.
- Рецепторную, реагируя на внешние раздражители. Подобную функцию осуществляют белки (гормоны, нейромедиаторы), которые получают сигналы-воздействия извне. Это служит отправной точкой для изменения хода обменных процессов внутри клетки.
- Ферментативную или метаболическую, когда мембранные белки-ферменты участвуют в разных химических процессах метаболизма. Большинство ферментов связаны с мембраной и в липидной оболочке созданы определенные условия для их работы, поэтому плазмалемма непосредственно и косвенно влияет на процессы метаболизма.
- Энергообразующую, которую осуществляют митохондрии – органоиды, которые относят к «маленьким силовым станциям клетки». Многие процессы, связанные с обменом между клеткой и межклеточным пространством нуждаются в дополнительных затратах энергии. Клетки также способны обмениваться энергией. Этот процесс осуществляется через белковые каналы.
- Матричную, когда цитолемма определяет местонахождение, фиксирует расположение органоидов внутри клетки и их позицию относительно друг друга. Оптимальное размещение позволяет органоидам легче взаимодействовать друг с другом.
- Маркировочную, так как каждая клетка снабжена «этикеткой» или маркировкой. Это антигены, состоящие из гликопротеинов (белков с разветвленными олигосахаридными цепями). Разветвления имеют разные конфигурации, поэтому каждый тип клеток снабжен уникальным «ярлыком», что позволяет идентифицировать или узнавать их. Например, иммунные клетки человека (макрофаги) распознают инородную клетку, проникшую в организм (бактерию или вирус). Они начинают работу по ее уничтожению. Таким же путем организм избавляется от старых, больных, мутировавших клеток.
Плазмалемма или мембрана – важнейшая составная часть живой клетки. Она выполняет не меньше функций, чем любой другой органоид. Имеет сложное строение и таит в себе много загадок, которые еще не раскрыты учеными. Этот «живой пропускной шлюз» продолжают изучать, чтобы использовать на благо человека.
Источники изображений:
- Рис. 1. — www.youtube.com/watch?v=uaC8KhzuH98&ab_channel=BogdanTuziak
- Рис. 3. — uchitel.pro/строение-и-функции-клетки/
- Рис. 4. — 900igr.net
Узнаем как ие функции выполняет наружная клеточная мембрана? Строение наружной клеточной мембраны
Изучением строения клеток прокариотических организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и клеточная стенка, а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция — это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.
Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная клеточная мембрана, строение и функции которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать плазмалемму клетки-хозяина для построения свой защитной оболочки.
А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в цитоплазму клетки и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех эукариотических клеток систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
Главные химические соединения, которые её образуют — это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.
Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы – структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в межклеточных контактах. У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.
Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы – распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.
Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе пассивного транспорта энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. Активный транспорт происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.
Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.
Внешняя мембрана — обзор
3.1 OMV: состав, биогенез и функциональные роли
OMV представляют собой устойчивые и дискретные сферические двухслойные липидные наноструктуры диаметром от 10 до 300 нм, происходящие из оболочки клетки и неспособные к независимой репликации ( Kulp, Kuehn, 2010; Huang et al., 2016) (рис.4). Отделение OMV было впервые обнаружено более 50 лет назад на микрографах, сделанных с помощью просвечивающего электронного микроскопа, на которых изображена ультраструктура клеточной стенки бактерий (Bladen and Waters, 1963; Bayer and Anderson, 1965; Chatterjee and Das, 1967).Было установлено, что эти наносферические структуры представляют собой единую мембрану, окружающую электронно-плотный центр (Work et al., 1966). Другие исследования того же периода также сообщили о наличии «внеклеточных глобул» в бесклеточном супернатанте E . coli культивировали в условиях ограничения роста питательных веществ (Bishop and Work, 1965; Knox et al., 1966; Work et al., 1966). Поэтому изначально предполагалось, что образование OMV происходит исключительно в стрессовых условиях.Тем не менее, дальнейшие исследования ясно показали, что OMV также можно обнаружить в нестрессовых условиях, как в лабораторных условиях, так и в условиях окружающей среды (Hoekstra et al., 1976; Hellman et al., 2000). В настоящее время известно, что OMV вносят вклад в множество ключевых биологических функций, и одна из первых ролей, когда-либо описанных для OMV, — это их участие в патогенезе, особенно в качестве средств доставки факторов вирулентности (обзор Ellis and Kuehn, 2010). С годами OMV были приписаны дополнительные функции, в зависимости от вида и условий культивирования, включая внутри- и межвидовое общение, реакцию на стрессы оболочки, приобретение питательных веществ, горизонтальный перенос генов, действия в качестве агентов-приманок, а также общественные блага ( подробное описание см. в Kulp and Kuehn, 2010 и Schwechheimer and Kuehn, 2015).В целом, OMV, по-видимому, способствуют выживанию бактерий в определенной экологической нише, что подчеркивает их значимость для бактериального гомеостаза.
Рис. 4. Везикулы наружной мембраны грамотрицательных бактерий. (A) Изображение везикулы наружной мембраны (OMV — , верхняя панель ), полученная из оболочки бактериальной клетки (нижняя панель) . Показаны детали внешней мембраны (OM) и внутренней мембраны (IM), включая трансмембранные белки. Периплазматическое пространство, в котором находится слой пептидогликана (PG), показывает растворимые периплазматические белки, неправильно свернутые белки и нуклеиновые кислоты.Содержание OMV иллюстрирует часть биомолекул, которые были идентифицированы как на их мембране, так и в просвете. (B и C) Электронные микрофотографии в просвете уранилацетата, отрицательно окрашенные Synechocystis sp. Клетка PCC 6803, высвобождающая OMV (B, увеличение 120 000 ×), и бесклеточная концентрированная внеклеточная среда Synechocystis sp. PCC 6803, показывающий несколько OMV (C, увеличение 40000).
(A) По данным января, A.T., 2017. Везикулы наружной мембраны (OMV) грамотрицательных бактерий: перспективное обновление.Фронт. Microbiol. 8, 1053.Многочисленные исследования показали, что OMV обогащены компонентами OM, а именно LPS и OMP, а также периплазматическими белками, фрагментами PG и даже цитоплазматическими и нуклеиновыми кислотами (Biller et al., 2014, 2017; Lee et al. ., 2016). В ранних сообщениях фактически не дифференцировали MV, искусственно сформированные в растворе (из-за естественного липидного поведения, заключающегося в перегруппировке в пузырьки, неизбирательно захватывая материал от бактериального лизиса) от интактных OMV. Совсем недавно улучшенные методологии изоляции и современные омические методы позволили провести тщательный анализ состава OMV.Примечательно, что OMV на самом деле обогащены определенными клеточными компонентами и обеднены другими (Lee et al., 2008), что подтверждает идею о том, что выбор содержимого груза не является случайным процессом. Например, Salmonella sp. Содержание OMV варьировалось в зависимости от тестируемых условий роста: в OMV, выделенных из клеток в условиях обилия питательных веществ, преимущественно выявлялись цитозольные белки, участвующие в трансляции и клеточном метаболизме, в то время как в ограниченных условиях питания OMV были обогащены мембранными белками, участвующими в транспорте питательных веществ (Bai и другие., 2014). Кроме того, подход, основанный на масс-спектрометрии, показал, что в OMV не было обнаружено широко консервативного специфического компонента (Schwechheimer et al., 2013), что еще раз указывает на переменный состав. В целом, ожидается, что дифференциальные составы OMV связаны как с зависимыми от штамма особенностями клеточной оболочки, так и с отдельными экологическими нишами (Yoon, 2016).
Было предложено три не исключающих друг друга механизма образования OMV. В одной модели везикуляция происходит, когда ковалентные поперечные связи между мембранными белками и слоем PG локально разрываются либо за счет временного уменьшения общего количества поперечных связей, либо за счет локального смещения поперечных связей, способствуя выпучиванию небольших ОМ порциями.Другая модель включает периплазматические нанотерритории, в которых накапливаются неправильно свернутые белки и другие компоненты оболочки (фрагменты LPS или PG). После этого аномального, ограниченного скопления клеточных компонентов целостность оболочки локально снижается, вызывая образование пузырей в частях ОМ, загруженных содержимым просвета. Наконец, также было высказано предположение, что определенные биофизические свойства некоторых липидов OM могут способствовать везикуляции за счет точного определения специфической интеграции LPS или фосфолипидов, что приводит к изменениям текучести и гибкости мембран.Также предполагается, что многие другие факторы влияют на размер, скорость продукции и состав OMV, и, если существует консенсусный процесс биогенеза OMV, он не полностью охарактеризован (Kulp, Kuehn, 2010; Schwechheimer and Kuehn, 2015; Yoon, 2016). ).
В исследованиях цианобактерий область OMV еще совсем недавно, и многое еще предстоит изучить. Это особенно хорошо иллюстрируется тем фактом, что самая ранняя публикация, посвященная исключительно изучению цианобактериальных OMV, датируется 2014 годом (Biller et al., 2014). В этом новаторском исследовании не только показано, что лабораторно контролируемые культуры морской цианобактерии Prochlorococcus постоянно выделяют OMV, но также и то, что эти везикулы можно найти в большом количестве в пробах морской воды. Кроме того, было продемонстрировано, что OMV Prochlorococcus были способны поддерживать рост гетеротрофных бактериальных культур, участвуя в этих структурах в морских потоках углерода. Кроме того, наблюдались взаимодействия морских фагов и везикул, показывающие способность OMV действовать как «ловушки».В целом авторы проиллюстрировали некоторые фундаментальные роли OMV и их бесчисленные значения для микробных экосистем (Biller et al., 2014). В более поздней публикации OMV Prochlorococcus сравнивали с таковыми трех других морских гетеротрофов в попытке раскрыть частоту упаковки ДНК в везикулы и различия между различными таксонами (Biller et al., 2017). Путем изучения количества и распределения ДНК, связанной с OMV, было показано, что ДНК по-разному инкапсулирована внутри популяций OMV и между ними.Более того, эта работа предполагает, что механизм упаковки ДНК в OMV не работает одинаково у всех бактерий (Biller et al., 2017). Помимо Prochlorococcus и морских Synechococcus было показано, что другие цианобактерии также образуют и высвобождают OMV, включая одноклеточный Synechococcus sp. PCC 7002 (Xu et al., 2013) и Synechocystis sp. PCC 6803 (Pardo et al., 2015; Oliveira et al., 2016), нитевидный Jaaginema litorale LEGE 07176 (Brito et al., 2017), а также нитевидная, образующая гетероцисты Anabaena sp. PCC 7120 (Oliveira et al., 2015a) и Cylindrospermopsis raciborskii (CYRF-01) (Zarantonello et al., 2018).
Помимо функций, описанных выше для OMV, происходящих из морских цианобактерий (Biller et al., 2014), для этих внеклеточных везикул были предложены другие функции. Высвобождение OMV цианобактериями может работать как эффективный путь секреции. Метаболически модифицированный Synechococcus sp.Было показано, что штамм PCC 7002, лишенный двух генов гликогенсинтазы, glgA -I и glgA -II, выделяет значительно больше OMV, чем штамм дикого типа (Xu et al., 2013). Авторы предположили, что, поскольку этот мутант экспортирует спонтанно растворимые сахара в среду, наблюдаемые OMV могут быть связаны с этим механизмом секреции, даже несмотря на то, что содержание сахара в наблюдаемых OMV не оценивалось (Xu et al., 2013) . Кроме того, Synechocystis sp.Штамм PCC 6803, лишенный гомолога TolC (необходимого для мембранно-зависимых механизмов секреции; см. Рис. 1 и 3), также продемонстрировал высвобождение значительно большего количества OMV, чем родительский штамм (Oliveira et al., 2016). Поскольку нокаут tolC был сильно нарушен в секреции внутриклеточных белков, метаболитов и экзогенных соединений, было высказано предположение, что гипервезикуляция может удовлетворить потребность в секреции. В согласии с этим, цианобактериальные OMV также были предложены для транспортировки материала, необходимого для развития биопленок.Это было предложено при наблюдении везикул, происходящих от цианобионтов, в спорокарпе водного папоротника Azolla microphylla (Zheng et al., 2009). Более того, поскольку генетический материал, как сообщается, наблюдался внутри этих пузырьков, они могли быть векторами для латерального переноса генов между цианобионтом и папоротником (Zheng et al., 2009). Однако цианобактериальные OMV могут также работать как механизм для снятия стресса оболочки: Gonçalves et al. охарактеризован набор из Synechocystis sp. Штаммы PCC 6803, лишенные нескольких компонентов транслоказы IM, участвующих в TolC-зависимых системах секреции (Gonçalves et al., 2018). Интересно отметить, что среди различных штаммов, обладающих разной способностью к высвобождению OMV, нокаут tolC (самый высокий продуцент OMV в исследовании) был единственным, демонстрирующим удивительно высокие уровни транскриптов spy и degQ , кодирующих белки, участвующие в стрессовые реакции оболочки и сверхэкспрессия Spy и DegP (Gonçalves et al., 2018). Таким образом, авторы предположили, что делеция tolC вызывает стресс оболочки, и что гипервезикуляция при нокауте tolC представляет собой независимый механизм борьбы с такими стрессовыми состояниями (Gonçalves et al., 2018).
Клеточные стенки прокариот | Безграничная микробиология
Клеточная стенка бактерий
Бактерии защищены жесткой клеточной стенкой, состоящей из пептидогликанов.
Цели обучения
Напомним характеристики клеточной стенки бактерий
Ключевые выводы
Ключевые моменты
- Клеточная стенка — это слой, расположенный за пределами клеточной мембраны растений, грибов, бактерий, водорослей и архей.
- Клеточная стенка пептидогликана, состоящая из дисахаридов и аминокислот, обеспечивает структурную поддержку бактерий.
- Стенка бактериальной клетки часто является мишенью для лечения антибиотиками.
Ключевые термины
- бинарное деление : Процесс, при котором клетка делится бесполым путем с образованием двух дочерних клеток.
Стенка бактериальной клетки : Анатомия структуры бактериальной клетки.
Бактериальные клетки не имеют ядра, связанного с мембраной.Их генетический материал обнажен внутри цитоплазмы. Рибосомы — их единственный тип органелл. Термин «нуклеоид» относится к области цитоплазмы, где расположена хромосомная ДНК, обычно к единственной кольцевой хромосоме. Бактерии обычно одноклеточные, за исключением случаев, когда они существуют в колониях. Эти предковые клетки воспроизводятся посредством бинарного деления, дублируя свой генетический материал, а затем по существу расщепляясь, образуя две дочерние клетки, идентичные родительской. Стенка, расположенная за пределами клеточной мембраны, обеспечивает клеточную поддержку и защиту от механического воздействия или повреждения в результате осмотического разрыва и лизиса.Основным компонентом бактериальной клеточной стенки является пептидогликан или муреин. Эта жесткая структура пептидогликана, специфичная только для прокариот, придает форму клетке и окружает цитоплазматическую мембрану. Пептидогликан — это огромный полимер дисахаридов (гликанов), сшитых короткими цепями мономеров идентичных аминокислот (пептидов). Костяк молекулы пептидогликана состоит из двух производных глюкозы: N-ацетилглюкозамина (NAG) и N-ацетилмурамовой кислоты (NAM) с пентапептидом, выходящим из NAM, который незначительно варьируется среди бактерий.Нити NAG и NAM синтезируются в цитозоле бактерий. Они связаны межпептидными мостиками. Они переносятся через цитоплазматическую мембрану с помощью молекулы-носителя, называемой бактопренолом. От пептидогликана внутрь все бактериальные клетки очень похожи. В дальнейшем мир бактерий делится на два основных класса: грамположительные (грамм +) и грамотрицательные (грамм -). Клеточная стенка обеспечивает важные лиганды для адгезии и рецепторы для вирусов или антибиотиков.
Грамотрицательная внешняя мембрана
Стенка грамотрицательных клеток состоит из внешней мембраны, слоя пептидоглигана и периплазмы.
Цели обучения
Распознавать характеристики грамотрицательных бактерий
Ключевые выводы
Ключевые моменты
- Наружная мембрана грамотрицательных бактерий содержит липополисахариды, белки и фосфолипиды.
- Липополисахаридный компонент действует как фактор вирулентности и вызывает заболевания у животных.
- Больше факторов вирулентности скрывается в периплазматическом пространстве между внешней мембраной и плазматической мембраной.
Ключевые термины
- липополисахарид : любой из большого класса липидов, конъюгированных с полисахаридами
- эндотоксин : Любой токсин, выделяемый микроорганизмом и высвобождающийся в окружающую среду только после его смерти.
Структура грамотрицательной клеточной стенки : грамотрицательная внешняя мембрана, состоящая из липополисахаридов.
У грамотрицательных бактерий клеточная стенка состоит из одного слоя пептидогликана, окруженного мембранной структурой, называемой внешней мембраной. Грамотрицательные бактерии не сохраняют кристаллический фиолетовый, но способны сохранять контрастное пятно, обычно сафранин, который добавляется после кристаллического фиолетового. Сафранин отвечает за красный или розовый цвет, который наблюдается у грамотрицательных бактерий. Клеточная стенка грамотрицательных бактерий тоньше (10 нанометров) и менее компактна, чем у грамположительных бактерий, но остается прочной, жесткой и эластичной, что придает им форму и защищает от экстремальных условий окружающей среды.Наружная мембрана грамотрицательных бактерий неизменно содержит уникальный компонент, липополисахарид (ЛПС) в дополнение к белкам и фосфолипидам. Молекула LPS токсична и классифицируется как эндотоксин, который вызывает сильный иммунный ответ, когда бактерии заражают животных.
У грамотрицательных бактерий внешняя мембрана обычно рассматривается как часть внешнего листка мембранной структуры и относительно проницаема. Он содержит структуры, которые помогают бактериям прикрепляться к животным клеткам и вызывать болезни.Слой пептидогликана нековалентно прикреплен к молекулам липопротеинов, называемым липопротеинами Брауна, через их гидрофобную головку. Между внешней мембраной и плазматической мембраной находится концентрированный гелеобразный матрикс (периплазма) в периплазматическом пространстве. Фактически, он является неотъемлемой частью грамотрицательной клеточной стенки и содержит белки, связывающие аминокислоты, сахара, витамины, железо и ферменты, необходимые для питания бактерий. Периплазматическое пространство может действовать как резервуар для факторов вирулентности и динамического потока макромолекул, представляющих метаболический статус клетки и ее реакцию на факторы окружающей среды.Вместе плазматическая мембрана и клеточная стенка (внешняя мембрана, слой пептидогликана и периплазма) составляют грамотрицательную оболочку.
Оболочка грамположительных клеток
Уграмположительных бактерий клеточная оболочка состоит из толстого слоя пептидогликанов.
Цели обучения
Сравните и сравните грамположительные и отрицательные пятна
Ключевые выводы
Ключевые моменты
- Грамположительные бактерии окрашиваются в фиолетовый цвет при окрашивании по Граму из-за присутствия пептидогликана в их клеточной стенке.
- Пептидогликаны прикрепляются к отрицательно заряженным мономерам липотейхоевой кислоты, важным для направления и прикрепления клеток.
- Липотейхоевые кислоты ковалентно связаны с липидами внутри цитоплазматической мембраны, таким образом связывая пептидогликаны с цитоплазмой клетки.
Ключевые термины
- Окраска по Граму : метод дифференциации видов бактерий на две большие группы (грамположительные и грамотрицательные).
Грамположительные бактерии окрашиваются по Граму в темно-синий или фиолетовый цвет.Хотя окрашивание по Граму является ценным диагностическим инструментом как в клинических, так и в исследовательских целях, не все бактерии могут быть окончательно классифицированы с помощью этого метода, что позволяет сформировать группы с грамм-переменной и грамм-неопределенной группой.
Грамположительные бактерии : Эти бактерии окрашивают в фиолетовый цвет при окрашивании по Граму.
Он основан на химических и физических свойствах их клеточных стенок. В первую очередь, он обнаруживает пептидогликан, который присутствует в толстом слое у грамположительных бактерий.Грамположительный результат имеет пурпурный / синий цвет, а грамотрицательный — розовый / красный цвет. Окрашивание по Граму почти всегда является первым шагом в идентификации бактериального организма и является окрашиванием по умолчанию, выполняемым лабораториями над образцом, когда не указывается конкретная культура.
У грамположительных бактерий клеточная стенка толстая (15-80 нанометров) и состоит из нескольких слоев пептидогликана. У них отсутствует внешняя мембранная оболочка, характерная для грамотрицательных бактерий. Перпендикулярно слоям пептидогликана проходит группа молекул, называемых тейхоевыми кислотами, которые уникальны для грамположительной клеточной стенки.Тейхоевые кислоты представляют собой линейные полимеры полиглицерина или полирибита, замещенные фосфатами и некоторыми аминокислотами и сахарами.
Полимеры тейхоевой кислоты иногда прикрепляются к плазматической мембране (называемой липотейхоевой кислотой, LTA) и, по-видимому, направлены наружу под прямым углом к слоям пептидогликана. Тейхоевые кислоты придают грамположительной клеточной стенке общий отрицательный заряд из-за наличия фосфодиэфирных связей между мономерами тейхоевой кислоты. Функции тейхоевой кислоты полностью не изучены, но считается, что она служит хелатирующим агентом и средством прилипания бактерий.Они необходимы для жизнеспособности грамположительных бактерий в окружающей среде и обеспечивают химическую и физическую защиту.
Одна из идей состоит в том, что они обеспечивают канал для регулярно ориентированных отрицательных зарядов для прохождения положительно заряженных веществ через сложную сеть пептидогликанов. Другая теория заключается в том, что тейхоевые кислоты каким-то образом участвуют в регуляции и сборке субъединиц мурамовой кислоты на внешней стороне плазматической мембраны.
Есть случаи, особенно в стрептококках, где тейхоевые кислоты участвуют в прикреплении бактерий к тканевым поверхностям и, как считается, вносят свой вклад в патогенность грамположительных бактерий.
Микоплазмы и другие бактерии с дефицитом клеточной стенки
У некоторых бактерий отсутствует клеточная стенка, но они сохраняют способность к выживанию, живя внутри другой клетки-хозяина.
Цели обучения
Различать бактерии с клеточными стенками и без них
Ключевые выводы
Ключевые моменты
- Примерами бактерий, у которых отсутствует клеточная стенка, являются микоплазмы и бактерии L-формы.
- Микоплазма является важной причиной болезней животных и не подвержена влиянию антибиотиков, направленных на синтез клеточной стенки.
- Микоплазмы получают холестерин из окружающей среды и образуют стерины для построения своей цитоплазматической мембраны.
Ключевые термины
- осмотическая среда : среда с контролируемым чистым перемещением молекул из области с высокой концентрацией растворителя в область с низкой концентрацией растворителя через проницаемую мембрану.
Для большинства бактериальных клеток клеточная стенка имеет решающее значение для выживания клеток, но есть некоторые бактерии, у которых нет клеточных стенок. Виды Mycoplasma являются широко распространенными примерами, и некоторые из них могут быть внутриклеточными патогенами, которые растут внутри своих хозяев. Такой бактериальный образ жизни называется паразитарным или сапрофитным. Клеточные стенки здесь не нужны, потому что клетки живут только в контролируемой осмотической среде других клеток. Вполне вероятно, что в какой-то момент в прошлом у них была способность формировать клеточную стенку, но поскольку их образ жизни превратился в существование внутри других клеток, они потеряли способность формировать стенки.
Бактерии L-формы : Бактерии L-формы не имеют структуры клеточной стенки.
В соответствии с этим очень ограниченным образом жизни внутри других клеток, эти микробы также имеют очень маленькие геномы. Им не нужны гены для всех видов биосинтетических ферментов, поскольку они могут украсть у хозяина последние компоненты этих путей. Точно так же им не нужны гены, кодирующие множество различных путей для различных источников углерода, азота и энергии, поскольку их внутриклеточная среда полностью предсказуема. Из-за отсутствия клеточных стенок Mycoplasma имеют сферическую форму и быстро погибают при помещении в среду с очень высокой или очень низкой концентрацией соли.Однако у Mycoplasma действительно есть необычно прочные мембраны, которые более устойчивы к разрыву, чем другие бактерии, поскольку этой клеточной мембране приходится бороться с факторами клетки-хозяина. Присутствие стеринов в мембране способствует их долговечности, помогая увеличить силы, удерживающие мембрану вместе. Другие виды бактерий иногда мутируют или реагируют на экстремальные условия питания, образуя клетки без стенок, называемые L-формами. Это явление наблюдается как у грамположительных, так и у грамотрицательных видов.L-формы имеют разнообразную форму и чувствительны к осмотическому шоку.
Клеточные стенки архей
Стенки клеток архей отличаются от стенок бактериальных клеток своим химическим составом и отсутствием пептидогликанов.
Цели обучения
Укажите сходство между клеточными стенками архей и бактерий
Ключевые выводы
Ключевые моменты
- Археи — одноклеточные микроорганизмы, у которых отсутствует клеточное ядро и связанные с мембраной органеллы.
- Как и другие живые организмы, археи имеют полужесткую клеточную стенку, защищающую их от окружающей среды.
- Клеточная стенка архей состоит из S-слоев и лишена молекул пептидогликана, за исключением метанобактерий, у которых в клеточной стенке есть псевдопептидогликан.
Ключевые термины
- целлюлоза : сложный углевод, который образует основную составляющую клеточной стенки большинства растений и играет важную роль в производстве многих продуктов, таких как бумага, текстиль, фармацевтические препараты и взрывчатые вещества.
- хитин : сложный полисахарид, полимер N-ацетилглюкозамина, обнаруженный в экзоскелетах членистоногих и в клеточных стенках грибов; считается ответственным за некоторые формы астмы у людей.
- цитоплазма : содержимое клетки, кроме ядра. Он включает цитозоль, органеллы, везикулы и цитоскелет.
Как и другие живые организмы, клетки архей имеют внешнюю клеточную мембрану, которая служит защитным барьером между клеткой и окружающей средой.Внутри мембраны находится цитоплазма, где выполняются жизненные функции археона и где расположена ДНК. Вокруг почти всех клеток архей находится клеточная стенка, полужесткий слой, который помогает клетке сохранять свою форму и химическое равновесие. Все эти три области можно выделить в клетках бактерий и большинства других живых организмов.
Археи : скопление галобактерий (архей)
Более пристальный взгляд на каждую область показывает структурное сходство, но значительные различия в химическом составе между бактериальной и архейной клеточной стенкой.Археи строят те же структуры, что и другие организмы, но строят их из разных химических компонентов. Например, клеточные стенки всех бактерий содержат химический пептидогликан. Стенки клеток архей не содержат это соединение, хотя некоторые виды содержат подобное. Он состоит из белков поверхностного слоя, называемых S-слоями. Точно так же археи не производят стенки из целлюлозы (как растения) или хитина (как грибы). Клеточная стенка архей химически различна. Единственным исключением являются метаногены, которые содержат в своей клеточной стенке цепи псевдопептидогликана, в химическом составе которых отсутствуют аминокислоты и N-ацетилмурамовая кислота.Наиболее разительные химические различия между археями и другими живыми существами заключаются в их клеточных мембранах. Существует четыре фундаментальных различия между мембраной архей и мембран всех других клеток: (1) хиральность глицерина, (2) эфирная связь, (3) изопреноидные цепи и (4) разветвление боковых цепей.
Повреждение клеточной стенки
Клеточная стенка отвечает за выживание бактериальных клеток и защиту от факторов окружающей среды и антимикробного стресса.
Цели обучения
Обсудить влияние повреждения клеточной стенки на бактериальную клетку
Ключевые выводы
Ключевые моменты
- Грамположительные и грамотрицательные бактерии защищены внешней клеточной стенкой, состоящей из различных слоев пептидогликана.
- Повреждение стенки бактериальной клетки нарушает ее целостность и создает дисбаланс электролитов, вызывающий гибель клетки.
- Некоторые классы антибиотиков действуют путем ингибирования синтеза строительных блоков клеточной стенки, что приводит к лизису и гибели клеток.
Ключевые термины
- гидролаза : фермент, катализирующий гидролиз субстрата.
- транспептидаза : любой фермент, катализирующий перенос амино- или пептидной группы от одной молекулы к другой
Клеточная стенка является основным элементом, несущим стресс и сохраняющим форму у бактерий.Таким образом, его целостность имеет решающее значение для жизнеспособности конкретной клетки. Как у грамположительных, так и у грамотрицательных бактерий каркас клеточной стенки состоит из поперечно-сшитого полимерного пептидогликана. Клеточная стенка грамотрицательных бактерий тонкая (всего около 10 нанометров в толщину) и обычно состоит всего из двух-пяти слоев пептидогликана, в зависимости от стадии роста. У грамположительных бактерий клеточная стенка намного толще (от 20 до 40 нанометров).
В то время как пептидогликан обеспечивает структурную основу клеточной стенки, считается, что тейхоевые кислоты, составляющие примерно 50% материала клеточной стенки, контролируют общий поверхностный заряд стенки.Это влияет на активность муреингидролазы, устойчивость к антибактериальным пептидам и адгезию к поверхностям. Хотя обе эти молекулы полимеризуются на поверхности цитоплазматической мембраны, их предшественники собираются в цитоплазме. Любое событие, которое мешает сборке предшественника пептидогликана и транспорту этого объекта через клеточную мембрану, где он интегрируется в клеточную стенку, может поставить под угрозу целостность стенки. Повреждение клеточной стенки нарушает состояние электролитов клетки, что может активировать пути гибели (апоптоз или запрограммированная гибель клеток).Регулируемая гибель и лизис клеток у бактерий играет важную роль в определенных процессах развития, таких как компетентность и развитие биопленок. Они также играют важную роль в устранении поврежденных клеток, например, необратимо поврежденных окружающей средой или антибиотическим стрессом. Примером антибиотика, нарушающего синтез клеточной стенки бактерий, является пенициллин. Пенициллин действует путем связывания с транспептидазами и ингибирования перекрестного связывания субъединиц пептидогликана. Бактериальная клетка с поврежденной клеточной стенкой не может подвергнуться бинарному делению и поэтому неизбежно погибнет.
Механизм действия пенициллина : Пенициллин действует путем связывания с пенициллин-связывающими белками и ингибирования перекрестного связывания субъединиц пептидогликана.
Структура и функции бактериальных клеток
Структура и функции бактериальных клеток (стр. 6)(В этой главе 10 страниц)
© Кеннет Тодар, доктор философии
Наружная мембрана Грамотрицательные бактерии ,00
Представляет особый интерес как компонент Грамотрицательный клеточная стенка внешняя мембрана , дискретная двухслойная структура на внешней стороне листа пептидогликана (см. рисунок 18 ниже).Для в бактерия, внешняя мембрана — это прежде всего проницаемость барьер но в первую очередь из-за содержания липополисахаридов он обладает многими интересные и важные характеристики грамотрицательных бактерий. В внешняя мембрана представляет собой липидный бислой, интеркалированный белками, поверхностно напоминающий плазматическую мембрану.Внутренняя поверхность внешней мембраны состоит из фосфолипидов, аналогичных фосфоглицеридам, которые составляют плазматическая мембрана. Наружная поверхность внешней мембраны может содержать некоторые фосфолипид, но в основном он образован другим типом амфифильный молекула, состоящая из липополисахарида (ЛПС). Внешняя мембрана белки обычно проходят через мембрану и в одном случае закрепляют внешний мембрана нижележащего пептидогликанового листа.
Рисунок 18. Схема иллюстрация внешней мембраны, клеточной стенки и плазматической мембраны грамотрицательных бактерия. Обратите внимание на структуру и расположение молекул, которые составлять внешняя мембрана.
Молекула ЛПС, которая составляет внешний лицо внешней мембраны состоит из гидрофобной области, называемой липидом . A , который присоединен к области гидрофильного линейного полисахарида, состоящий из основного полисахарида и O-специфичного полисахарид .
Рисунок 19. Структура LPS
Липид A глава молекула вставляется в внутренняя часть мембраны и полисахаридный хвост молекула сталкивается с водной средой. Куда вставляется хвост молекулы в в голове происходит накопление отрицательных зарядов, так что магний катион хелатируется между соседними молекулами ЛПС.Это обеспечивает боковой стабильность внешней мембраны и объясняет, почему лечение Грамотрицательный бактерии с мощным хелатирующим агентом, таким как ЭДТА, вызывают разброс молекул ЛПС.
Бактериальный липополисахариды токсичны для животные. При введении в небольших количествах LPS или эндотоксин активирует макрофаги для производства пирогенов, активирует каскад комплемента, вызывая воспаление, и активирует факторы крови, приводящие к внутрисосудистому свертыванию и кровоизлияние.Эндотоксины могут играть роль в заражении любым Грамотрицательный бактерия. Токсичным компонентом эндотоксина (ЛПС) является липид А. O-специфический полисахарид может предоставлять лиганды для прикрепления бактерий и некоторая устойчивость к фагоцитозу. Различия в точном содержании сахара из полисахарид O (также называемый антигеном O) составляет несколько антигенные типы (серотипы) среди грамотрицательных бактериальных патогенов.Следовательно. хотя липид А является токсичным компонентом ЛПС, полисахариды тем не менее способствуют вирулентности грамотрицательных бактерий.
Белки в внешняя мембрана Escherichia coli хорошо охарактеризованы (см. Таблицу 5). Около 400,00 экземпляров в Браун липопротеин ковалентно присоединен к листу пептидогликана на один конец и вставлен в гидрофобную внутреннюю часть мембраны на противоположный конец.Группа тримерных белков под названием поринов образует поры фиксированный диаметр через липидный бислой мембраны. В omp C и omp F поринов E. coli разработаны, чтобы позволить проход гидрофильных молекул до mw около 750 дальтон. Больше молекулы или вредные гидрофобные соединения (например, соли желчных кислот в кишечнике тракт) исключены из записи.Порины разработаны в грамотрицательных бактерии чтобы позволить прохождение полезных молекул (питательных веществ) через барьер наружную мембрану, но для исключения проникновения вредных веществ из окружающая обстановка. Повсеместно распространенный белок omp A во внешней мембране из E. coli имеет пориновую структуру и может функционировать в усвоение специфических ионов, но он также является рецептором пилуса F и вложение сайт бактериальных вирусов.
Стол 5. Функции внешний Компоненты мембраны Escherichia coli .
| Компонент | Функция |
| Липополисахарид (ЛПС) | Барьер проницаемости |
| Мосты Mg ++ | Стабилизирует LPS и важен для его проницаемость характеристики |
| Липопротеин Брауна | Закрепляет внешнюю мембрану на пептидогликане (мурейн) лист |
| Порины Omp C и Omp F | белков, которые образуют поры или каналы через внешняя мембрана для прохождения гидрофильных молекул |
| Omp A белок | обеспечивает рецептор для некоторых вирусов и бактериоцины; стабилизирует спаривающиеся клетки при конъюгации |
S-слои
белков S-слоя образуют самый внешний компонент оболочки клетки широкий спектр бактерий и архей.S-слои состоят из не замужем белка или гликопротеина (Mw 40-200 кДа) и проявляют либо косая, квадратная или гексагональная симметрия решетки с размерами элементарной ячейки в диапазоне от 3 до 30 нм. S-слои обычно имеют толщину от 5 до 10 нм и имеют поры одинакового размера (диаметр от 2 до 8 нм) и морфологии.
Кристаллические белки поверхностного слоя (S-слоя) бактериальных клеток были оптимизированы в течение миллиардов лет биологической эволюции как составляющие элементы одной из простейших систем самосборки в природа.Изолированные белки S-слоя обладают внутренним свойством: перекристаллизоваться в двумерные массивы в широком спектре поверхности, включая кремний, металлы и полимеры, а также интерфейсы, такие как плоские липидные пленки и липосомы. Четко определенное расположение функциональные группы на решетках S-слоя позволяют связывать молекулы и частицы в определенных регулярных массивах. S-слои также представляют шаблоны для формирования неорганических нанокристаллических сверхрешеток состоит из CdS, Au, Ni, Pt или Pd.
Самостоятельная сборка S-слоев иллюстрирует основной принцип строительства в природа для создания больших массивов биомолекул с четко определенными геометрические и физико-химические свойства поверхности.
Многие Грамотрицательные и грамположительные бактерии, а также многие археи имеют регулярно структурированный слой, называемый S-слоем, прикрепленный к крайняя часть их клеточной стенки.Он состоит из белка или гликопротеина и на электронных микрофотографиях имеет рисунок, напоминающий кафельная поверхность. Просвечивающая электронная микрофотография замороженного, металлически затененный препарат бактериальной клетки с S-слоем с шестиугольная решетка симметрия. Бар = 100 нм.S-уровни связаны с рядом возможных функций. В S-слой может защитить бактерии от вредных ферментов или изменений pH.Это может способствовать вирулентности, защищая бактерия против атаки комплемента и фагоцитоза. Считается защитить кишечную палочку от нападения хищной бактерией Bdellovibrio.
S-слой может действовать как адгезин, позволяя бактериям прилипать к мембранам клеток-хозяев и поверхностям окружающей среды, чтобы колонизировать.Многие из связанных с клетками белков адгезинов, используемых патогены являются компонентами S-слоя.
Корреляция между Реакция окрашивания по Граму и Свойства клеточной стенки бактерий сведены в Таблицу 6. График пятно процедура содержит этап «обесцвечивания», на котором клетки промывают с смесь ацетон-спирт.Содержание липидов грамотрицательной стенки вероятно, влияет на результат этого шага, так что грамположительные клетки удерживать первичное окрашивание, в то время как грамотрицательные клетки обесцвечиваются.
Стол 6. Соотношение Граммы окраска с другими свойствами Бактерий.
| Имущество | Грамположительные | Грамотрицательные |
| Толщина стенки | толщиной (20-80 нм) | тонкий (10 нм) |
| Количество слоев | 1 | 2 |
| Содержание пептидогликана (муреина) | > 50% | 10-20% |
| Тейхоевые кислоты в стенке | настоящее время | отсутствует |
| Содержание липидов и липопротеинов | 0–3% | 58% |
| Содержание белка | 0 | 9% |
| Содержание липополисахаридов | 0 | 13% |
| Чувствительность к пенициллину G | да | № (1) |
| Чувствительность к лизоциму | да | № (2) |
(2) Грамотрицательные бактерии чувствительны к лизоциму при предварительной обработке какой-либо процедурой, удаляющей наружную мембрану и подвергает пептидогликан непосредственно ферменту. Формы без клеточной стенки
Некоторые бактерии способны жить или существовать без клеточная стенка. Микоплазмы — это группа бактерий, у которых отсутствует клетка. стена. Микоплазмы содержат стеролоподобные молекулы, включенные в их мембраны и обычно они обитатели осмотически защищенных сред. Микоплазма pneumoniae является причиной первичной атипичной бактериальной пневмонии, известный в просторечии как «ходячая пневмония». По понятным причинам пенициллин неэффективен при лечении этого типа пневмонии. Иногда под давление антибактериальной терапии, патогенные бактерии могут вернуться в формы без клеточной стенки (называемые сферопластами или протопластами ) и сохраняются или выживать в осмотически защищенных тканях.Когда антибиотик отменен терапии организмы могут отрастить свои клеточные стенки и повторно инфицировать незащищенный ткани.
продолжение главы
Предыдущая страница
© Кеннет Тодар, доктор философии.D. Все права защищены. — www.textbookofbacteriology.net
Строение и функция белковых комплексов митохондриальной мембраны | BMC Biology
Риццуто Р., Де Стефани Д., Рафаэлло А., Маммукари С. Митохондрии как сенсоры и регуляторы передачи сигналов кальция. Nat Rev Mol Cell Biol. 2012; 13 (9): 566–78.
CAS Статья PubMed Google Scholar
Пеллегрино MW, Хейнс CM. Митофагия и митохондриальный развернутый белковый ответ при нейродегенерации и бактериальной инфекции. BMC Biol. 2015; 13:22.
PubMed Central Статья PubMed Google Scholar
Chandel NS. Митохондрии как сигнальные органеллы. BMC Biol. 2014; 12:34.
PubMed Central Статья PubMed Google Scholar
Bratic A, Larsson NG. Роль митохондрий в старении. J Clin Invest. 2013. 123 (3): 951–7.
PubMed Central CAS Статья PubMed Google Scholar
Gray MW, Lang BF, Cedergren R, Golding GB, Lemieux C, Sankoff D, et al. Структура генома и содержание генов в митохондриальной ДНК протистов. Nucleic Acids Res. 1998. 26 (4): 865–78.
PubMed Central CAS Статья PubMed Google Scholar
Amunts A, Brown A, Bai X, Llacer JL, Hussain T., Emsley P, et al. Структура большой рибосомной субъединицы митохондрий дрожжей. Наука. 2014; 343 (6178): 1485–9.
PubMed Central CAS Статья PubMed Google Scholar
Шмидт О., Пфаннер Н., Мейзингер С. Импорт митохондриального белка: от протеомики к функциональным механизмам. Nat Rev Mol Cell Biol. 2010. 11 (9): 655–67.
CAS Статья PubMed Google Scholar
Lackner LL. Формирование динамической митохондриальной сети. BMC Biol. 2014; 12:35.
PubMed Central Статья PubMed Google Scholar
Detmer SA, Chan DC. Функции и нарушения митохондриальной динамики. Nat Rev Mol Cell Biol. 2007. 8 (11): 870–9.
CAS Статья PubMed Google Scholar
Хоппинс С., Лакнер Л., Нуннари Дж.Машины, которые делят и объединяют митохондрии. Анну Рев Биохим. 2007. 76: 751–80.
CAS Статья PubMed Google Scholar
Alexandre A, Reynafarje B, Lehninger AL. Стехиометрия векторных движений H +, связанных с транспортом электронов и синтезом АТФ в митохондриях. Proc Natl Acad Sci U S. A. 1978; 75 (11): 5296–300.
PubMed Central CAS Статья PubMed Google Scholar
Meeusen S, McCaffery JM, Nunnari J. Промежуточные продукты слияния митохондрий, обнаруженные in vitro. Наука. 2004; 305: 1747–52.
CAS Статья PubMed Google Scholar
Гутман М., Котляр А.Б., Боровок Н., Нахлиэль Э. Реакция объемных протонов с препаратом внутренней митохондриальной мембраны: измерения с временным разрешением и их анализ. Биохимия. 1993. 32 (12): 2942–6.
CAS Статья PubMed Google Scholar
Перкинс Г.А., Фрей Т.Г. Недавние исследования структуры митохондрий с помощью микроскопии. Микрон. 2000. 31 (1): 97–111.
CAS Статья PubMed Google Scholar
Liao M, Cao E, Julius D, Cheng Y. Структура ионного канала TRPV1, определенная с помощью электронной криомикроскопии. Природа. 2013; 504 (7478): 107–12.
PubMed Central CAS Статья PubMed Google Scholar
Кюльбрандт В. Революция в разрешении. Наука. 2014; 343 (6178): 1443–4.
Артикул PubMed Google Scholar
Дэвис К.М., Штраус М., Даум Б., Киф Дж. Х., Осевач HD, Рыцовска А. и др. Макромолекулярная организация АТФ-синтазы и комплекса I в целых митохондриях. Proc Natl Acad Sci U S. A. 2011; 108 (34): 14121–6.
PubMed Central CAS Статья PubMed Google Scholar
Бхарат Т.А., Дэйви Н.Э., Ульбрих П., Ричес Д.Д., де Марко А., Румлова М. и др. Структура незрелого ретровирусного капсида при разрешении 8 A с помощью криоэлектронной микроскопии. Природа. 2012. 487 (7407): 385–9.
CAS Статья PubMed Google Scholar
Байрхубер М., Майнс Т., Хабек М., Беккер С., Гиллер К., Виллингер С. и др. Структура человеческого потенциалзависимого анионного канала. Proc Natl Acad Sci U S. A. 2008; 105 (40): 15370–5.
PubMed Central CAS Статья PubMed Google Scholar
Ллопис Дж., Маккаффери Дж. М., Мияваки А., Фаркуар М. Г., Цзянь Р. Я. Измерение цитозольного, митохондриального и рН Гольджи в отдельных живых клетках с зелеными флуоресцентными белками. Proc Natl Acad Sci U S. A. 1998; 95 (12): 6803–8.
PubMed Central CAS Статья PubMed Google Scholar
Kukat C, Wurm CA, Spahr H, Falkenberg M, Larsson NG, Jakobs S. Микроскопия со сверхвысоким разрешением показывает, что митохондриальные нуклеоиды млекопитающих имеют одинаковый размер и часто содержат единственную копию мтДНК. Proc Natl Acad Sci U S. A. 2011; 108 (33): 13534–9.
PubMed Central CAS Статья PubMed Google Scholar
Кукат К., Дэвис К., Вурм К.А., Спер Х., Бонекамп Н.А., Кюль И. и др. Перекрестное связывание TFAM с одной молекулой мтДНК образует митохондриальный нуклеоид.Proc Natl Acad Sci U S A. 2015. В печати.
Pfeffer S, Woellhaf MW, Herrmann JM, Forster F. Организация митохондриального механизма трансляции, изученного in situ с помощью криоэлектронной томографии. Nat Commun. 2015; 6: 6019.
CAS Статья PubMed Google Scholar
Шлейер М., Нойперт В. Транспорт белков в митохондрии: промежуточные соединения транслокации, охватывающие места контакта между внешней и внутренней мембранами.Клетка. 1985. 43 (1): 339–50.
CAS Статья PubMed Google Scholar
Перкинс Г., Ренкен С., Мартоне М.Э., Янг С.Дж., Эллисман М., Фрей Т. Электронная томография митохондрий нейронов: трехмерная структура и организация крист и мембранных контактов. J. Struct Biol. 1997. 119 (3): 260–72.
CAS Статья PubMed Google Scholar
Gold VA, Ieva R, Walter A, Pfanner N, van der Laan M, Kuhlbrandt W. Визуализация активных мембранных белковых комплексов с помощью электронной криотомографии. Nat Commun. 2014; 5: 4129.
PubMed Central CAS Статья PubMed Google Scholar
Pebay-Peyroula E, Dahout-Gonzalez C, Kahn R, Trezeguet V, Lauquin GJ, Brandolin G. Структура митохондриального носителя АДФ / АТФ в комплексе с карбоксиатрактилоидом. Природа. 2003. 426 (6962): 39–44.
CAS Статья PubMed Google Scholar
Sjostrand FS. Электронная микроскопия митохондрий и двойных мембран цитоплазмы. Природа. 1953. 171 (4340): 30–2.
CAS Статья PubMed Google Scholar
Palade GE. Электронно-микроскопическое исследование структуры митохондрий. J Histochem Cytochem. 1953; 1 (4): 188–211.
CAS Статья PubMed Google Scholar
Дэвис К.М., Ансельми С., Виттиг И., Фаральдо-Гомес Дж. Д., Кюльбрандт В. Структура димера дрожжевой F1Fo-АТФ-синтазы и его роль в формировании митохондриальных крист. Proc Natl Acad Sci U S. A. 2012; 109 (34): 13602–7.
PubMed Central CAS Статья PubMed Google Scholar
Pfanner N, van der Laan M, Amati P, Capaldi RA, Caudy AA, Chacinska A, et al. Единая номенклатура для сайта контакта митохондрий и системы организации крист.J Cell Biol. 2014. 204 (7): 1083–6.
PubMed Central CAS Статья PubMed Google Scholar
von der Malsburg K, Muller JM, Bohnert M, Oeljeklaus S, Kwiatkowska P, Becker T, et al. Двойная роль митофилина в организации митохондриальной мембраны и биогенезе белков. Dev Cell. 2011. 21 (4): 694–707.
Артикул PubMed Google Scholar
Vogel F, Bornhovd C, Neupert W, Reichert AS. Динамическая субкомпартментализация внутренней мембраны митохондрий. J Cell Biol. 2006. 175 (2): 237–47.
PubMed Central CAS Статья PubMed Google Scholar
фон Баллмос С., Виденманн А., Димрот П. Основы синтеза АТФ с помощью АТФ-синтаз F1F0. Анну Рев Биохим. 2009. 78: 649–72.
Артикул Google Scholar
Вт IN, Монтгомери MG, Рансуик MJ, Leslie AG, Walker JE. Биоэнергетическая стоимость создания молекулы аденозинтрифосфата в митохондриях животных. Proc Natl Acad Sci U S. A. 2010; 107 (39): 16823–7.
PubMed Central CAS Статья PubMed Google Scholar
Stock D, Leslie AGW, Walker JE. Молекулярная архитектура вращательного двигателя в АТФ-синтазе. Наука. 1999; 286: 17701705.
Артикул Google Scholar
Allen RD, Schroeder CC, Fok AK. Исследование внутренних мембран митохондрий методами быстрого замораживания и глубокого травления. J Cell Biol. 1989. 108 (6): 2233–40.
CAS Статья PubMed Google Scholar
Strauss M, Hofhaus G, Schroder RR, Kühlbrandt W. Димерные ленты АТФ-синтазы формируют внутреннюю митохондриальную мембрану. EMBO J. 2008; 27 (7): 1154–60.
PubMed Central CAS Статья PubMed Google Scholar
Allegretti M, Klusch N, Mills DJ, Vonck J, Kühlbrandt W, Davies KM. Горизонтальные присущие мембране альфа-спирали в а-субъединице статора АТФ-синтазы F-типа. Природа. 2015; 521 (7551): 237–40.
CAS Статья PubMed Google Scholar
Кюльбрандт В., Карен Д. АТФ-синтазы: новый поворот для древней машины. Trends Biochem Sci. 2015. В печати.
Paumard P, Vaillier J, Coulary B, Schaeffer J, Soubannier V, Mueller DM, et al.АТФ-синтаза участвует в формировании морфологии митохондриальных крист. EMBO J. 2002; 21 (3): 221–30.
PubMed Central CAS Статья PubMed Google Scholar
Bornhövd C, Vogel F, Neupert W, Reichert AS. Потенциал митохондриальной мембраны зависит от олигомерного состояния супракомплексов F1F0-АТФ-синтазы. J Biol Chem. 2006. 281 (20): 13990–8.
Артикул PubMed Google Scholar
Джорджио В., фон Штокум С., Антониэль М., Фаббро А., Фоголари Ф., Форте М. и др. Димеры митохондриальной АТФ-синтазы образуют поры перехода проницаемости. Proc Natl Acad Sci U S. A. 2013; 110 (15): 5887–92.
PubMed Central CAS Статья PubMed Google Scholar
Hunte C, Koepke J, Lange C, Roßmanith T., Michel H. Структура комплекса цитохрома bc1 дрожжевого Saccharomyces cerevisiae, совместно кристаллизованного с Fv-фрагментом антитела, с разрешением 2,3 Å.Структура. 2000. 8: 669–84.
CAS Статья PubMed Google Scholar
Цукихара Т., Аояма Х., Ямасита Э., Томизаки Т., Ямагути Х., Синдзава-Ито К. и др. Вся структура 13-субъединицы окисленной цитохром с оксидазы при 2,8 А. Наука. 1996. 272 (5265): 1136–44.
CAS Статья PubMed Google Scholar
Цикерманн В., Вирт С., Насири Х., Зигмунд К., Швальбе Х., Хант С. и др.Механистическое понимание кристаллической структуры митохондриального комплекса I. Наука. 2015; 347 (6217): 44–9.
CAS Статья PubMed Google Scholar
Ефремов Р.Г., Барадаран Р., Сазанов Л.А. Архитектура дыхательного комплекса I. Природа. 2010. 465 (7297): 441–5.
CAS Статья PubMed Google Scholar
Виноткумар К.Р., Чжу Дж., Херст Дж.Архитектура респираторного комплекса млекопитающих I. Природа. 2014. 515 (7525): 80–4.
PubMed Central CAS Статья PubMed Google Scholar
Schägger H, Pfeiffer K. Суперкомплексы в дыхательных цепях митохондрий дрожжей и млекопитающих. EMBO J. 2000; 19 (8): 1777–83.
PubMed Central Статья PubMed Google Scholar
Schäfer E, Seelert H, Reifschneider NH, Krause F, Dencher NA, Vonck J.Архитектура активных суперкомплексов дыхательной цепи млекопитающих. J Biol Chem. 2006. 281 (22): 15370–5.
Артикул PubMed Google Scholar
Althoff T, Mills DJ, Popot JL, Kühlbrandt W. Расположение компонентов цепи переноса электронов в митохондриальном суперкомплексе крупного рогатого скота I1III2IV1. EMBO J. 2011; 30 (22): 4652–64.
PubMed Central CAS Статья PubMed Google Scholar
Lapuente-Brun E, Moreno-Loshuertos R, Acin-Perez R, Latorre-Pellicer A, Colas C, Balsa E, et al. Сборка суперкомплекса определяет поток электронов в митохондриальной цепи переноса электронов. Наука. 2013; 340 (6140): 1567–70.
CAS Статья PubMed Google Scholar
Mileykovskaya E, Penczek PA, Fang J, Mallampalli VK, Sparagna GC, Dowhan W. Расположение комплексов дыхательной цепи в суперкомплексе Saccharomyces cerevisiae III2IV2, выявленное с помощью криоэлектронной микроскопии одиночных частиц.J Biol Chem. 2012. 287 (27): 23095–103.
PubMed Central CAS Статья PubMed Google Scholar
Блаза Дж. Н., Серрели Р., Джонс А. Дж., Мохаммед К., Херст Дж. Кинетические доказательства против разделения пула убихинона и каталитической значимости суперкомплексов дыхательной цепи. Proc Natl Acad Sci U S. A. 2014; 111 (44): 15735–40.
PubMed Central CAS Статья PubMed Google Scholar
Li P, Nijhawan D, Budihardjo I., Srinivasula SM, Ahmad M, Alnemri ES, et al. Цитохром c и dATP-зависимое образование комплекса Apaf-1 / каспаза-9 запускает каскад апоптотических протеаз. Клетка. 1997. 91 (4): 479–89.
CAS Статья PubMed Google Scholar
Даум Б., Вальтер А., Хорст А., Осиевач HD, Кюльбрандт В. Возрастная диссоциация димеров АТФ-синтазы и потеря крист внутренней мембраны в митохондриях.Proc Natl Acad Sci U S. A. 2013; 110 (38): 15301–6.
PubMed Central CAS Статья PubMed Google Scholar
Наварро А., Боверис А. Система митохондриальной трансдукции энергии и процесс старения. Am J Physiol Cell Physiol. 2007; 292 (2): C670–86.
CAS Статья PubMed Google Scholar
Dröse S, Brandt U. Молекулярные механизмы производства супероксида дыхательной цепью митохондрий.Adv Exp Med Biol. 2012; 748: 145–69.
Артикул PubMed Google Scholar
Twig G, Ширихай О.С. Взаимодействие митохондриальной динамики и митофагии. Антиоксидный окислительно-восстановительный сигнал. 2011. 14 (10): 1939–51.
PubMed Central CAS Статья PubMed Google Scholar
Scheckhuber CQ, Erjavec N, Tinazli A, Hamann A, Nyström T, Osiewacz HD.Уменьшение деления митохондрий приводит к увеличению продолжительности жизни и пригодности двух моделей старения грибов. Nat Cell Biol. 2007. 9 (1): 99–105.
CAS Статья PubMed Google Scholar
Calvo SE, Mootha VK. Митохондриальный протеом и болезнь человека. Анну Рев Геномикс Хум Генет. 2010; 11: 25–44.
PubMed Central CAS Статья PubMed Google Scholar
Хендерсон Р. Возможности и ограничения нейтронов, электронов и рентгеновских лучей для микроскопии с атомным разрешением неокрашенных биологических молекул. Q Rev Biophys. 1995; 28: 171–93.
CAS Статья PubMed Google Scholar
Рис Д.М., Лесли А.Г., Уокер Дж. Э.. Структура внешней части мембраны АТФ-синтазы крупного рогатого скота. Proc Natl Acad Sci U S. A. 2009; 106 (51): 21597–601.
PubMed Central CAS Статья PubMed Google Scholar
Symersky J, Pagadala V, Osowski D, Krah A, Meier T., Faraldo-Gomez JD, et al. Структура c (10) кольца митохондриальной АТФ-синтазы дрожжей в открытой конформации. Nat Struct Mol Biol. 2012; 19 (5): 485–91. S481.
PubMed Central CAS Статья PubMed Google Scholar
| Филогенетика | Геология | Биохимия | Клеточная биология | Экология | История жизни | Зоология | Ботаника | Палеогеография | | |
амебоид — не имеющий определенной формы клетки, способный изменять форму. | |
amphiesma — Наружное покрытие динофлагеллаты, состоящее из нескольких слоев мембраны. | |
отверстие — Маленькое отверстие, например отверстие при испытании пеной. | |
бактериофаг — Вирус, который заражает и уничтожает бактериального хозяина. Однако некоторые фаги встраивают свою ДНК в ДНК своего хозяина и остаются бездействующими в течение длительного периода.По этой причине они стали незаменимыми инструментами генных инженеров. | |
капсид — Белковая «оболочка» свободной вирусной частицы. | |
ячейка — фундаментальная структурная единица всего живого. Клетка состоит в основном из внешней плазматической мембраны, которая отделяет ее от окружающей среды; генетический материал (ДНК), который кодирует наследуемую информацию для поддержания жизни; и цитоплазма, гетерогенная совокупность ионов, молекул и жидкости. | |
клеточный цикл — Полная последовательность шагов, которые должны быть выполнены клеткой для самовоспроизведения, как видно от митотического события к митотическому событию. Большая часть цикла состоит из периода роста, в течение которого клетка набирает массу и реплицирует свою ДНК. Остановка клеточного цикла — важная особенность воспроизводства многих организмов, включая человека. | |
клеточная мембрана — Наружная мембрана клетки, отделяющая ее от окружающей среды.Также называется плазматической мембраной или плазмалеммой. | |
клеточная стенка — Жесткая структура, отложенная вне клеточной мембраны. Растения известны своими клеточными стенками из целлюлозы, как и зеленые водоросли и некоторые простейшие, в то время как грибы имеют клеточные стенки из хитина. | |
хлоропласт — хлорофилл-содержащая пластида, обнаруженная в клетках водорослей и зеленых растений. | |
хромосома — линейный фрагмент эукариотической ДНК, часто связанный специализированными белками, известными как гистоны. | |
ценоцитарный — Состояние, при котором организм состоит из нитчатых клеток с большими центральными вакуолями, ядра которых не разделены на отдельные части. В результате получается длинная трубка, содержащая множество ядер со всей цитоплазмой на периферии. | |
колониальный — Состояние, при котором многие одноклеточные организмы живут вместе в несколько скоординированной группе. В отличие от настоящих многоклеточных организмов, отдельные клетки сохраняют свою индивидуальность и, как правило, свои собственные мембраны и клеточные стенки. | |
сократительная вакуоль — У многих протистов специализированная вакуоль с соответствующими каналами, предназначенная для сбора избыточной воды в клетке. Микротрубочки периодически сокращаются, чтобы вытеснить избыток воды из клетки, регулируя осмотический баланс клетки. | |
цитоплазма — Все содержимое клетки, включая плазматическую мембрану, но не включая ядро. | |
цитоскелет — Интегрированная система молекул внутри эукариотических клеток, которая придает им форму, внутреннюю пространственную организацию, подвижность и может способствовать общению с другими клетками и окружающей средой.Например, красные кровяные тельца были бы сферическими, а не плоскими, если бы не их цитоскелет. | |
дикариотический — Имеющий два разных и разных ядра на клетку; содержится в грибах. Дикариотическая особь называется дикарионом. | |
диплоид — наличие двух разных наборов хромосом в одном ядре каждой клетки. Большинство многоклеточных животных и растений диплоидны. Сравните с гаплоидом. | |
двойная мембрана — В митохондриях и пластидах есть двухслойная мембрана, которая окружает органеллу.Считается, что это результат эндосимбиоза, когда внешняя мембрана происходит от эукариотической клетки, а внутренняя мембрана принадлежит первоначальному прокариоту, который был «проглочен». | |
эндоплазматический ретикулум — (ER) сеть мембран в эукариотических клетках, которая помогает контролировать синтез белка и клеточную организацию. | |
эукариот — н. Организм, клетки которого имеют цитоскелеты для поддержки, а их ДНК содержится в ядре, отделенном от другого содержимого клетки; е.g., простейшие, растения, животные и грибы; эукариот — прил. | |
внеклеточный матрикс — (ECM) Область вне клеток многоклеточных животных, которая включает соединения, прикрепленные к плазматической мембране, а также растворенные вещества, привлеченные поверхностным зарядом клеток. ЕСМ функционирует как для удержания животных клеток вместе, так и для их защиты от окружающей среды. | |
глазное пятно — Светочувствительная органелла, обнаруженная во многих группах простейших и у некоторых многоклеточных животных. | |
филамент — Длинная цепочка белков, например, в волосах, мышцах или жгутиках. | |
fission — Деление одноклеточных организмов, особенно прокариот, у которых митоз не происходит. Также используется для обозначения митоза у некоторых одноклеточных грибов. | |
жгутик — n. Волосоподобная структура, прикрепленная к клетке, используется для передвижения у многих простейших и прокариот.Прокариотический жгутик отличается от эукариотического жгутика тем, что прокариотический жгутик представляет собой твердую единицу, состоящую в основном из белка флагеллина, тогда как эукариотический жгутик состоит из нескольких белковых нитей, связанных мембраной, и не содержит флагеллина. Жгутик эукариот иногда называют ундулиподиумом. | |
панцирь — Минеральный «скелет» диатомовой водоросли или другого одноклеточного организма. | |
Аппарат Гольджи — эукариотическая органелла, которая упаковывает клеточные продукты, такие как ферменты и гормоны, и координирует их транспорт за пределы клетки. | |
гаплоид — Наличие одного набора хромосом в ядре каждой клетки. Мхи, многие простейшие и грибы гаплоидны, как и некоторые насекомые, мохообразные и гаметы. всех организмов. Контраст с диплоидный. | |
haptonema — Колышковая структура, уникальная для Prymnesiophyta; его функция неизвестна. | |
lorica — Внешнее покрытие в форме вазы или чашки.Обнаружен во многих простейших, включая некоторых жгутиконосцев, инфузорий, хризофитов и хоанофлагеллят, а также в некоторых клетках животных. | |
лизосома — эукариотическая органелла, несущая пищеварительные ферменты. Лизосома сливается с вакуолярной мембраной, содержащей проглоченные частицы, на которые затем действуют ферменты. | |
мастигонема — Небольшие волосовидные нити, обнаруженные на «волосатом» жгутике Chromista. | |
мембрана — В биологии пограничный слой внутри или вокруг живой клетки или ткани. | |
мезокариотический — Ядерное состояние, уникальное для динофлагеллят, при котором хромосомы остаются постоянно конденсированными. | |
микротрубочек — Тип филамента в эукариотических клетках, состоящий из единиц протеина тубулина. Помимо других функций, это основной структурный компонент жгутика эукариот. | |
микроворсинок — Тонкие пальцевидные выступы на поверхности клетки, часто используемые для увеличения абсорбционной способности или для улавливания частиц пищи. «Воротник» хоанофлагеллят на самом деле состоит из близко расположенных микроворсинок. | |
митохондрия — Сложная органелла, обнаруженная у большинства эукариот; Считается, что они произошли от свободноживущих бактерий, которые установили симбиотические отношения с примитивными эукариотами.Митохондрии являются местом производства большей части энергии у большинства эукариот; им для работы необходим кислород. См .: двойная мембрана . | |
митоз — Процесс ядерного деления у эукариот. Это один шаг в цитокинезе или клеточном делении. БОЛЕЕ ?. | |
MTOC — (центр организации микротрубочек) MTOC представляют собой пучки белковых трубок, которые могут быть обнаружены в основании жгутика эукариот.У животных они также участвуют в создании массивов микротрубочек, которые разделяют хромосомы во время митоза. | |
многоклеточный — Любой организм, состоящий из множества клеток, называется многоклеточным. | |
нм — n. Единица измерения; одна миллионная (10 -9 ) метра. | |
ядерная мембрана — Двойная мембрана, которая окружает эукариотическое ядро.На его поверхности много пор, которые регулируют поток крупных соединений в ядро и из него. | |
нуклеоид — Область в прокариотах, где сконцентрирована ДНК. В отличие от ядра он не связан мембраной. | |
ядро — Органелла, связанная с мембраной, которая содержит ДНК в виде хромосом. Это место репликации ДНК и место синтеза РНК. | |
органелла — н.Мембраносвязанная структура в эукариотической клетке, которая разделяет клетку на области, которые выполняют различные клеточные функции, например, митохондрии, эндоплазматический ретикулум, лизосомы. | |
плазматическая мембрана — Наружная мембрана клетки, иногда называемая клеточной мембраной. Термин «плазматическая мембрана» чаще используется при обсуждении прокариот. | |
плазмида — Круговая петля ДНК у прокариот.Эукариотическая ДНК организована в хромосомы. | |
пластида — Любая из нескольких пигментированных цитоплазматических органелл, обнаруженных в клетках растений и другие организмы, выполняющие различные физиологические функции, такие как синтез и хранение пищи. | |
прокариот — Буквально «перед ядром», этот термин применяется ко всем бактериям и архее. Прокариотические клетки не имеют внутренних мембран или цитоскелета.Их ДНК круговая, а не линейная. | |
протоплазма — Все содержимое клетки, включая ядро. (см .: цитоплазма) | |
псевдоподии — Пальцевидные отростки из амебоидной клетки; буквально «ложные ноги». | |
повторяющихся последовательностей — Длина нуклеотидной последовательности, которая повторяется в тандемном кластере. | |
ретикулоподии — Длинные нитевидные псевдоподии, которые разветвляются и снова соединяются, образуя тонкую сеть.Они характерны для форам. | |
рибосома — (рибосомная РНК) | |
синцитик — см. Hexactinellida | |
test — n. Твердая оболочка, производимая некоторыми одноклеточными простейшими; может состоять из карбоната кальция, кремнезема или песчинок. | |
theca — Общий термин для любого жесткого внешнего покрытия одноклеточного протиста, обычно состоящего из сцепляющихся пластин.динофлагелляты и диатомовые водоросли являются примерами простейших с теками. | |
трансдукция — Вирусный перенос ДНК новому хозяину. | |
трихоциста — Органелла инфузорий и динофлагеллят, которая выделяет длинные нитчатые белки при повреждении клетки. Используется как защита от потенциальных хищников. | |
ультраструктура — Подробная структура образца, например, клетки , ткани или органа , что можно наблюдать только с помощью электронной микроскопии.Также называется тонкой структурой. В яичной скорлупе ультраструктура относится к трехмерному расположению минеральных кристаллов и органического вещества. Он описывается с точки зрения минералогии кальцита или арагонита и перехода между различными зонами организации внутри оболочки. Отчетливые зоны организации называются зонами ультраструктуры. | |
undulipodium — Еще один термин для обозначения жгутика эукариот. | |
вакуоль — Мембранно-связанное заполненное жидкостью пространство внутри ячейки.В большинстве растительных клеток есть одна большая вакуоль, заполняющая большую часть объема клетки. Некоторые бактериальные клетки содержат газовые вакуоли. | |
| Последнее обновление: 2009-11-12 | |
Ячейки: их структура и функции
Ячейки: их структура и функции Глава 4: Структура и функции ячеекАнималкулы и первые микроскопы
А. Ранние микроскописты
1.Галилей видел детали глаз насекомых двумя грубыми линзы.2. Роберт Гук наблюдал за пробкой в простые линзы, в которые он видел крошечные отсеки, которые он назвал ячейками (целлюлозы) .
3. Ван Левенгук увидел протистанов, сперму и бактерии с помощью своего линзы и микроскопы.
Б. Теория клетки
1. Шлейден (ботаник) и Шванн (зоолог): Считалось, что все растения и животные состоят из клеток.2. Вирхов: клетки происходят из уже существующих клеток.
C. Теория клетки: три обобщения:
1. Все организмы состоят из одной или нескольких клеток.I. Основные аспекты клеточной структуры и Функция2. Ячейка — это наименьшая единица, обладающая свойствами жизнь.
3. Непрерывность жизни проистекает непосредственно из роста и деление одиночных клеток.
А.Структурная организация клеток
1. Ячейка — это наименьший объект, который все еще сохраняет характеристики жизни.2. Все ячейки состоят из трех основных частей:
а. Плазматическая мембрана отделяет каждую клетку от окружающая среда, разрешает поток молекул через мембрану, и содержит рецепторы, которые могут влиять на клеточные виды деятельности.г. Область, содержащая ДНК, занимает часть интерьер.
г. Цитоплазма содержит мембраносвязанные компартменты. (кроме бактерий), частицы и волокна & endash; все залито в полужидком веществе.
3. Эукариотические клетки определяются наличием у них мембраносвязанное ядро.
4. Прокариотические клетки не имеют определенного ядра; единственный представители — бактерии.
B. Жидкая мозаичная модель клеточных мембран
1.«Жидкая» часть клеточной мембраны состоит из фосфолипиды.а. Молекула фосфолипида состоит из гидрофильная голова и два гидрофобных хвоста.г. Если молекулы фосфолипидов окружены водой, их кластер хвостов гидрофобных жирных кислот и двухслойный результат; гидрофильные головки находятся на внешних гранях двухслойной лист.
г. Бислои фосфолипидов — структурная основа для всех клеточных мембран.
2. Внутри бислоя фосфолипиды показывают довольно много движение; они рассеиваются в стороны, вращаются и сгибают хвосты, чтобы предотвращают плотную упаковку и способствуют текучести, что также приводит к от липидов с короткими хвостами и ненасыщенных хвостов (изгиб при двойном облигации).
C. Обзор мембранных белков
1. Множество различных белков встроены в двухслойным или расположенным на двух его поверхностях.II. Размер и форма ячейки2. Мембранные белки служат транспортными белками, рецепторами. белки, белки распознавания и белки адгезии.
A. Из-за их небольшого размера большинство ячеек можно увидеть только с помощью светового и электронного микроскопов.
B. Размер ячейки должен быть небольшим; помните соотношение поверхности к объему!
Слишком большая ячейка не может двигаться материалы в камеру и из нее.III. В центре внимания наука: Микроскопы: путь к Ячейки
IV. Определяющие особенности эукариотических клеток
A. Основные сотовые компоненты
1. Органеллы образуют отделенные части цитоплазма.2. Все эукариотических клеток содержат органеллы.
а. Ядро контролирует доступ к ДНК и позволяет легче упаковывать ДНК во время деления клеток.г. Эндоплазматический ретикулум (ER) модифицирует новые образует полипептидные цепи, а также участвует в липидном синтез.
г. Корпус Гольджи модифицирует, сортирует и отправляет белки; они также играют роль в синтезе липидов для секреция или внутреннее употребление.
г. Везикулы транспортируют материал между органеллами и функция внутриклеточного пищеварения.
e. Митохондрии — эффективные фабрики АТФ производство.
3. Клетки также содержат немембранные структуры:
г. Рибосомы , «свободные» или прикрепленные к мембранам, участвуют в сборке полипептидных цепей.г. Цитоскелет помогает определять форму клеток, внутренняя организация и движения.
4.Органеллы разделяют реакции по времени (позволяя правильная последовательность) и пространство (позволяющее несовместимые реакции на происходят в непосредственной близости).
B. Какие органеллы типичны для растений?
1. На рис. 4.7a показано расположение частей растительной клетки.2. Несмотря на то, что он обозначен как «типичный», ни одна диаграмма не может говорить для всех разновидностей растительных клеток.
C. Какие органеллы типичны для животных?
1.На рис. 4.7b показано расположение частей клеток животных.V. Ядро2. Несмотря на то, что он обозначен как «типичный», ни одна диаграмма не может говорить для всех разновидностей клеток животных.
3. Также обратите внимание на различия между растительными и животными клетками, особенно клеточная стенка и большая центральная вакуоль растения клетки.
A. Ядро изолирует ДНК, которая содержит код белка сборка из участков (рибосомы в цитоплазме), где белки будут быть собранным.
1. Локализация ДНК упрощает сортировку наследственные инструкции, когда приходит время делиться клетке.2. Мембранная граница ядра помогает контролировать обмен сигналами и веществами между ядром и цитоплазма.
B. Ядерная оболочка
1. Ядерная оболочка состоит из двух липидных бислои с порами.2.Он окружает внутри нуклеоплазму.
3. На внутренней поверхности расположены места прикрепления белка. нити, которые закрепляют молекулы ДНК и удерживают их организовано.
C. Ядрышко
1. Ядрышко, расположенное внутри ядра, выглядит как более темная шаровидная масса.2. Это область, в которой субъединицы рибосом изготавливаются заранее. перед отгрузкой из ядра.
Д.Хромосомы
1. Хроматин относится к общему количеству клеток. сбор ДНК и связанных белков.2. Хромосома — это отдельная молекула ДНК и ее ассоциированные белки.
3. ДНК дублируется и конденсируется перед делением клетки. имеет место.
E. Что происходит с белками, определяемыми ДНК?
1. Некоторые из полипептидных цепей, собранных на рибосомы накапливаются в цитоплазме.VI. Цитомембранная система2. Другие проходят через цитомембранную систему, где они принимают в их окончательной форме и упаковываются в пузырьки для использования в ячейка или на экспорт.
A. Эндоплазматическая сеть
1. Эндоплазматический ретикулум представляет собой совокупность соединенные между собой трубки и сплюснутые мешочки, которые начинаются от ядра и побродить по цитоплазме.2.Различают два типа по наличию или отсутствию рибосом:
а. Rough ER состоит из уложенных друг на друга плоских мешочков. с множеством прикрепленных рибосом; олигосахаридные группы прикрепляются к полипептидам, когда они проходят через другие органеллы или секреторные пузырьки.г. Smooth ER не содержит рибосом; это район от какие везикулы, несущие белки и липиды, отпочковываются; это также инактивирует вредные химические вещества.
Тела Б. Гольджи
1. В теле Гольджи , белки и липиды проходят финальную обработку, сортировку и упаковку.2. Мембраны Гольджи уложены стопками из уплощенные мешочки, края которых разрываются в виде пузырьков.
C. Разнообразие пузырьков
1. Лизосомы — это пузырьки, которые отпочковываются от Гольджи. тела; они содержат мощные ферменты, которые могут переваривать содержимое других пузырьков, изношенных частей клеток или бактерий и чужеродных частицы.VII. Митохондрии2. Пероксисомы — это везикулы, содержащие ферменты, которые расщепляют жирные кислоты и аминокислоты; перекись водорода высвобожденный разрушается другим ферментом.
A. Митохондрии являются первичными органеллами для переноса энергия углеводов в АТФ в условиях избытка кислорода.
B. В клетках встречаются сотни тысяч митохондрий.
1.Имеет две мембраны, внутреннюю складчатую мембрану. (cristae) окружены гладкой внешней мембраной.VIII. Органеллы специализированные растения2. Внутренние и внешние отсеки, образованные мембранами, являются важно в преобразованиях энергии.
3. Митохондрии имеют собственную ДНК и некоторые рибосомы, факт. что указывает на возможность того, что когда-то они были независимыми сущности.
A. Хлоропласты и пластиды прочие
1. Хлоропласты имеют овальную или дискообразную форму, ограничены двойной мембраной и имеет решающее значение для процесса фотосинтез.а. В уложенных дисках (грана) пигменты и ферменты улавливают энергию солнечного света с образованием АТФ.г. Сахар образуется в жидком веществе (строме). окружающие стопки.
г. Пигменты, такие как хлорофилл (зеленый), придают отличительный цвета к хлоропластам.
2. Хромопласты содержат каротиноиды, которые Цвета частей растений от красного до желтого, но без хлорофилла.
3. Амилопласты не имеют пигментов; они хранят крахмал зерна в частях растений, таких как клубни картофеля.
B. Центральная вакуоль
1. В зрелом растении центральная вакуоль может занимают от 50 до 90% внутреннего пространства клетки!IX. Цитоскелета.хранит аминокислоты, сахар, ионы и отходы.г. увеличивается во время роста и значительно увеличивает площадь внешней поверхности клетки.
2. Цитоплазма вдавливается в очень узкую зону между центральная вакуоль и плазматическая мембрана.
A. Основные компоненты
1. Цитоскелет — это взаимосвязанная система волокна, нити и решетки, которые простираются между ядром и плазматическая мембрана.2. Придает клеткам их внутреннюю организацию, общую форму, и способность двигаться.
3. Основными компонентами являются микротрубочек, микрофиламентов и промежуточные филаменты: , все собраны из белка субъединицы.
4. Некоторые участки временные, например «шпиндель» микротрубочки, используемые для перемещения хромосом во время деления клеток; другие являются постоянными, например, волокна, работающие в мышцах. сокращение.
Б. Структурная основа движения клеток
1. За счет контролируемой сборки и разборки их субъединицы, микротрубочки и микрофиламенты растут или сжимаются длина (пример: движение хромосом).2. Микрофиламенты или микротрубочки активно скользят мимо них. другой (пример: движение мышц).
3. Микротрубочки или микрофиламенты шунтируют органеллы из одной местоположение в другое (пример: цитоплазматический поток).
C. Жгутики и реснички
1. Жгутики довольно длинные, обычно нет. многочисленны и обнаруживаются на одноклеточных простейших и сперматозоидах животных. клетки.X. Специализации клеточной поверхности2. Реснички короче и многочисленнее и могут обеспечить движение к свободноживущим клеткам или может перемещать окружающую воду и частицы, если ресничная клетка заякорена.
3. Оба этих расширения плазматической мембраны имеют 9 + 2 поперечный массив (возникающий из центриолей) и полезен в двигательная установка.
A. Стенки эукариотических клеток
1. Многие одноклеточные эукариоты имеют клеточную стенку, поддерживающая и защитная структура за пределами плазматической мембраны2. Микроскопические поры позволяют воде и растворенным веществам проходить к и от нижележащая плазматическая мембрана.
3. У растений пучки целлюлозных нитей образуют первичный клеточная стенка, которая более податлива, чем более жесткая вторичная стена, заложенная в ней позже.
4. Плазмодесматы — каналы, пересекающие соседние стенки. для соединения цитоплазмы соседних клеток.
B. Матрицы между клетками животных
1. Это сеть, которая удерживает клетки и ткани животных. вместе и влияет на то, как клетки будут делиться и метаболизировать.2. Хрящ состоит из клеток и белков (коллагена и эластин), рассеянный в основном веществе (модифицированный полисахариды).
C. Соединения между ячейками
1. На поверхности ткани клетки соединяются вместе, образуя барьер между интерьером и экстерьером.XI. Прокариотические клетки: бактерии2. Обычно используются три перехода от ячейки к ячейке.
а. Плотные соединения связывают клетки эпителия ткани для формирования уплотнений.г. Приклеивание стыков похоже на точечную сварку тканей подвержены растяжению.
г. Разрыв соединений связывает цитоплазму соседних клетки; они образуют каналы связи.
A. Термин прокариот (буквально «перед ядром») указывает на существование бактерий до эволюции клеток с ядро; бактериальная ДНК сгруппирована в отдельной области цитоплазма.
B. Бактерии — одни из самых маленьких и простых клеток.
1. Несколько жесткая клеточная стенка поддерживает клетку и окружает плазматическую мембрану, которая регулирует транспорт внутрь и из клетки.2. Рибосомы, сайты сборки белков, рассредоточены по всему телу. цитоплазма.
3. Жгутики бактерий (без массива 9 + 2) обеспечивают движение; пили на поверхности клетки помогают бактериям прикрепляться к поверхностям и Другой.
Клеточная мембрана — Определение и примеры
Определение
существительное
множественное число: клеточные мембраны
ˈmembɹeɪn
Наружная мембрана клетки состоит из двух слоев фосфолипидов со встроенными белками и отделяет содержимое клетки от ее внешняя среда, а также регулирует, что входит и выходит из клетки
Детали
Обзор
Клетка является структурной, функциональной и биологической единицей всех организмов.Это мембраносвязанная структура, содержащая цитоплазму и цитоплазматические структуры. Мембрана, которая окружает клетку и отделяет ее от внешней среды, называется клеточной мембраной . У животных плазматическая мембрана является самым внешним покрытием клетки, тогда как у растений, грибов и некоторых бактерий она расположена под клеточной стенкой. Хотя некоторые клетки образуют другой слой над клеточной мембраной (называемый клеточной стенкой), у других клеток клеточная мембрана является единственным защитным барьером между цитоплазмой и внешней стороной клетки.
Характеристики
Модель жидкой мозаики — теоретическая модель определенных биологических мембран, включая клеточную мембрану. Основываясь на этой модели, клеточная мембрана представляет собой липидный бислой фосфолипидов со встроенными белками и избирательно проницаема . Это означает, что некоторые вещества могут проходить через него (например, через пассивный транспорт). Другие вещества не смогли бы проходить через него без использования транспортных механизмов , таких как белков-носителей .Это жизненно важно для функции биологических мембран, которая заключается в регулировании того, что входит и выходит через них. Благодаря этой особенности клеточной мембраны она способна регулировать приток и отток веществ и тем самым помогает поддерживать гомеостаз. Более того, модель жидкой мозаики изображает клеточную мембрану , жидкость . Текучесть обеспечивается гидрофобными составными компонентами, такими как липиды и мембранные белки, которые перемещаются в поперечном или боковом направлении через мембрану. Это означает, что мембрана не твердая, а больше похожа на «жидкость».
Состав
Основными составляющими клеточной мембраны являются: липиды, белки и углеводы. Липиды бывают трех типов: фосфолипиды, гликолипиды и стерины. Фосфолипиды — это основной тип липидов в клеточной мембране. Это амфипатические соединения; «голова» гидрофильна, тогда как липофильный «хвост» гидрофобна.
Будучи амфипатическими, хвосты фосфолипидов имеют тенденцию избегать взаимодействия с водой. Напротив, головки фосфолипидов могут взаимодействовать с водой.Таким образом, при помещении в воду или водный раствор фосфолипиды имеют тенденцию к агрегированию, ориентируясь своими хвостами друг к другу. Таким образом, «головки» обычно обращены к воде или водному раствору. В результате фосфолипиды в клеточной мембране образуют характерный «липидный бислой».
Два слоя фосфолипидов расположены таким образом, что их гидрофобные хвосты выступают внутрь, а их гидрофильные головки выступают наружу. Такая организация фосфолипидов в клеточных мембранах делает последние избирательно проницаемыми для ионов и молекул.
Гликолипид — это углевод, ковалентно связанный с липидом. Гликолипиды — это биомолекулярные структуры в фосфолипидном бислое клеточной мембраны, углеводный компонент которых распространяется за пределы клетки. Другими липидами, содержание которых относительно невелико, являются гликолипиды и стерины. Гликолипиды (например, гликосфинголипид) необходимы для обеспечения стабильности и межклеточных взаимодействий, например клеточная адгезия с образованием ткани. Они также способствуют клеточному распознаванию, что важно для иммунологических функций.Стерол (например, холестерин) — это тип липидов в клеточной мембране, который обеспечивает структурную целостность и текучесть.
Холестерин в мембранах клеток животных позволяет клеткам животных изменять форму и поэтому они более гибкие, чем клетки растений (которые менее гибки по форме из-за наличия клеточной стенки). Из-за холестерина животным клеткам не обязательно иметь клеточные стенки, такие как стенки бактериальных и растительных клеток.
Белки — еще один важный компонент клеточной мембраны.Обычно они составляют около половины объема мембраны. Мембранные белки бывают трех основных типов: интегральных белков , периферических мембранных белков и липид-заякоренных белков . Интегральные белки — это мембранные белки, прочно прикрепленные к клеточной мембране.
Примерами являются трансмембранные белки (то есть белки, которые охватывают липидный бислой мембраны) и интегральные монотопные белки (то есть белки, которые постоянно прикреплены к мембране с одной стороны).Белки периферической мембраны — это белки, которые временно прикрепляются к мембране либо к липидному бислою, либо к интегральным белкам за счет комбинации гидрофобных, электростатических и других нековалентных взаимодействий. Заякоренные в липидах белки — это белки на поверхности клетки, которые ковалентно прикреплены к липидам в клеточной мембране.
Углеводы в клеточной мембране — это преимущественно гликопротеины. Гликопротеин относится к любому белку, ковалентно присоединенному к углеводной единице в процессе гликозилирования.Гликопротеины необходимы для распознавания клеток у эукариот.
Общие биологические реакции
Распознавание клеток — один из способов, с помощью которых клетки общаются друг с другом. Это возможно благодаря специфическим клеточным молекулам адгезии на поверхности клетки. Примером распознавания клеток является интегрин (LFA-1) связывания Т-клеток с ICAM эндотелиальной клетки. Другой — это селектин (L) связывания лимфоцитов с адресином (CD34) эндотелиальной клетки.
Одна из основных функций клеточной мембраны — транспортная.Клеточная мембрана участвует как в пассивном, так и в активном типе транспорта. При пассивном переносе вещества движутся по градиенту концентрации. Это контрастирует с активным переносом, который представляет собой тип транспорта, характеризующийся восходящим движением веществ (то есть от низшего к высшему) и, следовательно, требует химической энергии, например АТФ. При перемещении веществ через биологическую мембрану пассивный транспорт может нуждаться или не нуждаться в помощи мембранного белка.
Существует четыре основных типа пассивного транспорта: (1) простая диффузия, (2) облегченная диффузия, (3) фильтрация и (4) осмос.Простая и облегченная диффузия относится к чистому перемещению молекул от более высоких концентраций к более низким. Осмос относится к диффузии растворителя (обычно молекул воды) через полупроницаемую мембрану от более низких до более высоких концентраций растворенных веществ. Фильтрация — это движение молекул воды и растворенных веществ через клеточную мембрану под действием гидростатического давления, создаваемого сердечно-сосудистой системой.
Эндоцитоз — это процесс, при котором клетка поглощает материалы (например, белки и гормоны) извне, поглощая их и сливаясь со своей плазматической мембраной.Существует два типа эндоцитоза: фагоцитоз, что буквально означает поедание клеток, и пиноцитоз, что буквально означает потребление клеток.
Клетка поглощает , создавая небольшую деформацию внутрь (инвагинацию), содержащую вещество, которое должно транспортироваться внутри клетки. Затем инвагинация отделяется от клеточной мембраны, в результате чего образуется пузырек, содержащий вещество. Поскольку для эндоцитоза требуется АТФ, он считается одной из форм активного транспорта.
Экзоцитоз — это процесс, при котором клетка, кажется, выплевывает из клетки материалов.Таким образом, экзоцитоз кажется процессом, противоположным эндоцитозу. Везикула, содержащая материал, сливается с клеточной мембраной, а затем содержимое экструдируется за пределы клетки в окружающую среду.
Биологические функции
Структура и состав клеточной мембраны делают ее избирательно проницаемой (или полупроницаемой), что означает, что не каждому веществу разрешено проникать в клетку или покидать ее. Клеточная мембрана контролирует, какие вещества могут входить и выходить из клетки.Он может позволить определенному веществу пройти через определенное время, а затем отвергнуть то же самое вещество в более позднее время. Наличие поверхностных молекул (например, гликопротеинов, гликолипидов и т. Д.) Служит «сигнатурой» клетки. У каждой клетки есть своя «подпись» или «маркер», которые, как считается, работают при распознавании клеток или в своего рода системе клеточной идентификации. Его другие основные функции включают адгезию клеток, проводимость ионных каналов, передачу сигналов от клеток и точку прикрепления цитоскелета (что важно для сохранения формы клетки).
Дополнительный
Синоним (ы)
- плазматическая мембрана
- клеточная мембрана
- цитоплазматическая мембрана
- плазмалемма
Дополнительная литература
См. Также
© Biology Online. Контент предоставлен и модерируется Biology Online Editors
.