Что такое плазматическая мембрана, какого ее значения?
Плазматическая мембрана, или плазмалемма, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, ограничивающая клетку снаружи, что обусловливает ее непосредственную связь с внеклеточной средой, а следовательно, со всеми веществами и стимулами, воздействующими на клетку. Поэтому плазматическая мембрана играет роль барьера, преграды между сложно организованным внутриклеточным содержимым и внешней средой. В этом случае плазмалемма выполняет не только роль механического барьера, но, главное, ограничивает свободный поток низко- и высокомолекулярных веществ в обе стороны через мембрану. Более того, плазмалемма выступает как структура, «узнающая» , рецептирующая, различные химические вещества и регулирующая избирательно транспорт этих веществ в клетку и из нее. Другими словами, плазматическая мембрана осуществляет функции, связанные с регулируемым избирательным трансмембранным транспортом веществ, и исполняет роль первичного клеточного анализатора. В этом отношении плазмалемму можно считать клеточным органоидом, входящим в вакуолярную систему клетки. Как и другие мембраны этой системы (мембраны лизосом, эндосом, аппарата Гольджи и др.) , она возникает и обновляется за счет синтетической активности эндоплазматического ретикулума и имеет сходную композицию. Как ни странно, но плазматическую мембрану можно уподобить мембране внутриклеточной вакуоли, но вывернутой наизнанку: она не окружена гиалоплазмой, а окружает ее.
touch.otvet.mail.ru
Плазматическая мембрана Википедия
У этого термина существуют и другие значения, см. Мембрана. Модель клеточной мембраны. Маленькие голубые и белые шарики — гидрофильные «головки» фосфолипидов, а присоединённые к ним линии — гидрофобные «хвосты». На рисунке показаны только интегральные мембранные белки (красные глобулы и жёлтые спирали). Жёлтые овальные точки внутри мембраны — молекулы холестерина. Жёлто-зелёные цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликсОсновные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») части. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7—8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погружённые одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
История исследования
В 1925 году Гортер и Грендель с помощью осмотического «удара» получили так называемые «тени» эритроцитов — их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем с помощью ацетона выделили из оболочек липиды и определили количество липидов на единицу площади эритроцита — этого количества хватило на сплошной двойной слой. Хотя этот эксперимент привёл исследователей к правильному выводу, ими было допущено несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными плёнками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение — белки. В 1935 году Даниэлли и Доусон представили научному сообществу модель «сендвича», которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трёхслойное строение всех клеточных мембран.
Но постепенно накапливались аргументы против «бутербродной модели»:
- накапливались сведения о глобулярности плазматической мембраны;
- оказалось, что структура мембраны при электронной микроскопии зависит от способа её фиксации;
- плазматическая мембрана может различаться по структуре даже в одной клетке, например в головке, шейке и хвосте сперматозоида;
- «бутербродная» модель термодинамически не выгодна — для поддержания такой структуры нужно затрачивать большое количество энергии, и протащить вещество через мембрану очень сложно;
- количество белков, связанных с мембраной электростатически, очень небольшое, в основном белки очень тяжело выделить из мембраны, так как они погружены в неё.
Всё это привело к созданию в 1972 году С. Д. Сингером (S. Jonathan Singer) и Г. Л. Николсоном (Garth L. Nicolson) жидкостно-мозаичной модели строения мембраны. Согласно этой модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные, полуинтегральные и периферические. Периферические белки действительно находятся на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими взаимодействиями, но никогда не образуют сплошной слой. Доказательствами жидкостности мембраны служат методы FRAP, FLIP и соматическая гибридизация клеток, мозаичности — метод замораживания-скалывания, при котором на сколе мембраны видны бугорки и ямки, так как белки не расщепляются, а целиком отходят в один из слоёв мембраны.
Функции
- Барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой[1]. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- Транспортная — через мембрану происходит транспорт веществ в клетку и из клетки[1]. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортёры) и белки-каналы или путём эндоцитоза.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивает из неё ионы натрия (Na+). - Матричная — обеспечивает определённое взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- Механическая — обеспечивает автономность клетки, её внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечении механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- Энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки.
- Рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. - Ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- Осуществление генерации и проведения биопотенциалов.
- Маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединёнными к ним разветвлёнными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку.
Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются. Рядом с белками находятся аннулярные липиды — они более упорядочены, менее подвижны, имеют в составе более насыщенные жирные кислоты и выделяются из мембраны вместе с белком. Без аннулярных липидов белки мембраны не работают.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, в наружном содержатся преимущественно фосфатидилинозитол, фосфатидилхолин, сфингомиелины и гликолипиды, во внутреннем — фосфатидилсерин, фосфатидилэтаноламин и фосфатидилинозитол. Переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён, но может происходить спонтанно, примерно раз в 6 месяцев или с помощью белков-флиппаз и скрамблазы плазматической мембраны. Если в наружном слое появляется фосфатидилсерин, это является сигналом для макрофагов о необходимости уничтожения клетки.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
См. также
Примечания
- ↑ 1 2 Твердислов В. А., Яковенко Л. В. Физика биологических мембран // Школьникам о современной физике. Акустика. Теория относительности. Биофизика. — М., Просвещение, 1990. -ISBN 5-09-001323-3. — Тираж 200 000 экз. — С. 131-158
Литература
- Антонов В. Ф., Смирнова Е. Н., Шевченко Е. В. Липидные мембраны при фазовых переходах. — М.: Наука, 1994.
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). — 1-е издание. — М.: Мир, 1997. — ISBN 5-03-002419-0.
- Иванов В. Г., Берестовский Т. Н. Липидный бислой биологических мембран. — М.: Наука, 1982.
- Рубин А. Б. Биофизика, учебник в 2 тт. — 3-е издание, исправленное и дополненное. — М.: издательство Московского университета, 2004. — ISBN 5-211-06109-8.
- Bruce Alberts, et al. Molecular Biology Of The Cell. — 5th ed. — New York: Garland Science, 2007. — ISBN 0-8153-3218-1. — учебник по молекулярной биологии на английском языке
Ссылки
wikiredia.ru
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА. Функции. Структура
Оболочка клетки – это комплекс структур, отделяющий клетку от окружающей среды. Она состоит из наружного слоя – клеточной стенки и расположенной под ней плазматической мембраны.
Клетки животных и растений различаются по строению их наружного слоя. У растений и грибов на поверхности клеток расположена плотная оболочка — клеточная стенка. У большинства растений она состоит из целлюлозы, у грибов — из хитина. Клеточная стенка представляет собой защитную оболочку, обеспечивает форму растительных клеток, через клеточную стенку проходит вода, соли, молекулы многих органических веществ.
У животной клетки клеточной стенки нет. К цитоплазме примыкает плазматическая мембрана.
Под клеточной стенкой расположена плазматическая мембрана — плазмалемма (мембрана — кожица, пленка), граничащая непосредственно с цитоплазмой. Толщина плазматической мембраны около 10 нм.
Учитель: Сегодня на уроке мы познакомимся со строением и функциями плазматической мембраны.
Из истории открытия мембраны
Термин «мембрана» был предложен около ста лет назад для обозначения границ клетки, но с развитием электронной микроскопии стало ясно, что клеточная мембрана входит в состав структурных элементов клетки.
О наличии же пограничной мембраны между клеткой и окружающей средой было известно задолго до появления электронного микроскопа. Физико-химики отрицали существование плазматической мембраны и считали, что это просто граница раздела между живым коллоидным содержимым и окружающей средой, но Пфеффер (немецкий ботаник и физиолог растений) в 1890 году подтвердил существование ЦПМ.
В начале прошлого века Овертон (британский физиолог и биолог) обнаружил, что скорость проникновения многих веществ в эритроциты прямо пропорциональна их растворимости в липидах. В связи с этим ученый предположил, что мембрана содержит большое количество липидов и вещества, растворяясь в ней, проходят через нее и оказываются по ту сторону мембраны.
В 1925 году Гортер и Грендель (американские биологи) выделили липиды из клеточной мембраны эритроцитов. Полученные липиды они распределили по поверхности воды толщиной в одну молекулу. Оказалось, что площадь поверхности, занятой слоем липидов, в два раза больше площади самого эритроцита. Поэтому эти ученые сделали вывод, что клеточная мембрана состоит не из одного, а из двух слоев липидов.
Даусон и Даниэлли (английские биологи) в 1935 году высказали предположение, что в клеточных мембранах липидный бимолекулярный слой заключен между двумя слоями белковых молекул.
С появлением электронного микроскопа открылась возможность познакомиться со строением мембраны, и тогда обнаружилось, что мембраны животных и растительных клеток выглядят как трехслойная структура.
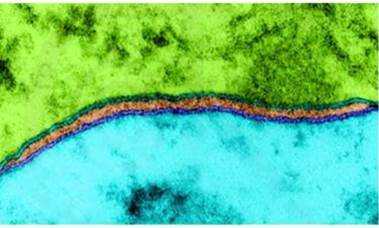
В 1959 году биолог Дж. Д. Робертсон, объединив имевшиеся в то время данные, выдвинул гипотезу о строении «элементарной мембраны», в которой он постулировал структуру, общую для всех биологических мембран.
Постулаты Робертсона о строении «элементарной мембраны»:
1. Все мембраны имеют толщину около 7,5 нм.
2. В электронном микроскопе все они представляются трехслойными.
3. Трехслойный вид мембраны есть результат именно того расположения белков и полярных липидов, которое предусматривала модель Даусона и Даниэлли – центральный липидный бислой заключен между двумя слоями белка.
Эта гипотеза о строении «элементарной мембраны» претерпела различные изменения и в 1972 году Сингером и Николсоном была предложена жидкостно-мозаичная модель мембраны, которая в настоящее время является общепризнанной.
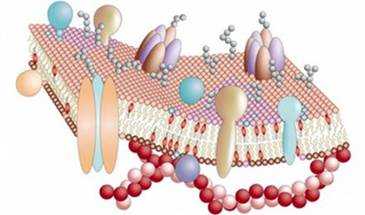
Согласно этой модели основой любой мембраны является двойной слой фосфолипидов. У фосфолипидов (соединений, содержащих фосфатную группу) молекулы состоят из полярной головки и двух неполярных хвостов.
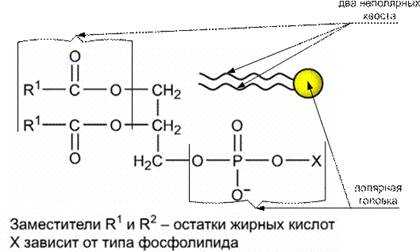
В фосфолипидном бислое гидрофобные остатки жирных кислот обращены внутрь, а гидрофильные головки, включающие остаток фосфорной кислоты, – наружу.
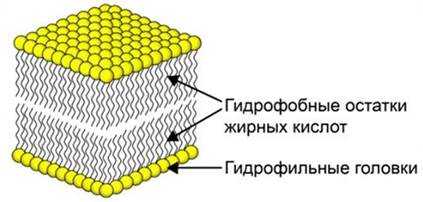
Фосфолипидный бислой представлен как динамическая структура, липиды могут перемещаться, меняя свое положение.
Двойной слой липидов обеспечивает барьерную функцию мембраны, не давая содержимому клетки растекаться, и препятствует попаданию в клетку токсических веществ.
Мембранные белки
В липидный бислой мембраны погружены молекулы белков, они образуют подвижную мозаику. По расположению в мембране и способу взаимодействия с липидным бислоем белки можно разделить на:
— поверхностные (или периферические) мембранные белки, связанные с гидрофильной поверхностью липидного бислоя;
— интегральные (мембранные) белки, погруженные в гидрофобную область бислоя.
Интегральные белки различаются по степени погруженности их в гидрофобную область бислоя. Они могут быть полностью погружены (интегральные) или частично погружены (полуинтегральные), а также могут пронизывать мембрану насквозь (трансмембранные).
Мембранные белки по своим функциям можно разделить на две группы:
— структурные белки. Они входят в состав клеточных мембран и участвуют в поддержании их структуры.
— динамические белки. Они находятся на мембранах и участвуют в происходящих на ней процессах.
Выделяют три класса динамических белков.
1. Рецепторные. С помощью этих белков клетка воспринимает различные воздействия на свою поверхность. То есть они специфически связывают такие соединения, как гормоны, нейромедиаторы, токсины на наружной стороне мембраны, что служит сигналом для изменения различных процессов внутри клетки или самой мембраны.
2. Транспортные. Эти белки транспортируют через мембрану те или иные вещества, также они образовывают каналы, через которые осуществляется транспорт различных ионов в клетку и из нее.
3. Ферментативные. Это белки-ферменты, которые находятся в мембране и участвуют в различных химических процессах.
На поверхности мембраны животных клеток образуется наружный рецепторный слой углеводов – гликокаликс. Образование гликокаликса, так же как и клеточных стенок растений, происходит благодаря жизнедеятельности самих клеток.
Основные функции клеточной мембраны
1. Структурная(клеточная мембрана отделяет клетку от окружающей среды).
2. Транспортная (через клеточную мембрану осуществляется транспорт веществ, причем клеточная мембрана является высокоизбирательным фильтром).
3. Рецепторная (находящиеся на поверхности мембраны рецепторы воспринимают внешние воздействия, передают эту информацию внутрь клетки, позволяя ей быстро реагировать на изменения окружающей среды).
Другие функции мембраны
Похожие статьи:
poznayka.org
Какое строение имеет плазматическая мембрана? Каковы ее функции?
Какое строение имеет плазматическая мембрана? Каковы ее функции?
Основу структурной организации клетки составляют биологические мембраны. Плазматическая мембрана (плазмалемма) — это мембрана, окружающая цитоплазму живой клетки. Мембраны состоят из липидов и белков. Липиды (в основном фосфолипиды) образуют двойной слой, в котором гидрофобные «хвосты» молекул обращены внутрь мембраны, а гидрофильные — к её поверхностям. Молекулы белков могут располагаться на внешней и внутренней поверхности мембраны, могут частично погружаться в слой липидов или пронизывать её насквозь. Большая часть погруженных белков мембран — ферменты. Это жидкостно-мозаичная модель строения плазматической мембраны. Молекулы белка и липидов подвижны, что обеспечивает динамичность мембраны. В состав мембран входят также углеводы в виде гликолипидов и гликопротеинов (гликокаликс), располагающихся на внешней поверхности мембраны. Набор белков и углеводов на поверхности мембраны каждой клетки специфичен и является своеобразным указателем типа клеток.
Функции мембраны:
- Разделительная. Она заключается в образовании барьера между внутренним содержимым клетки и внешней средой.
- Обеспечение обмена веществ между цитоплазмой и внешней средой. В клетку поступают вода, ионы, неорганические и органические молекулы (транспортная функция). Во внешнюю среду выводятся продукты, образованные в клетке (секреторная функция).
- Транспортная. Транспорт через мембрану может проходить разными путями. Пассивный транспорт осуществляется без затрат энергии, путем простой диффузии, осмоса или облегченной диффузии с помощью белков- переносчиков. Активный транспорт — с помощью белков- переносчиков, и он требует затрат энергии (например, натрий-калиевый насос).
Крупные молекулы биополимеров попадают внутрь клетки в результате эндоцитоза. Его разделяют на фагоцитоз и пиноцитоз. Фагоцитоз — захват и поглощение клеткой крупных частиц. Явление впервые было описано И.И. Мечниковым. Сначала вещества прилипают к плазматической мембране, к специфическим белкам-рецепторам, затем мембрана прогибается, образуя углубление.
Образуется пищеварительная вакуоль. В ней перевариваются поступившие в клетку вещества. У человека и животных к фагоцитозу способны лейкоциты. Лейкоциты поглощают бактерии и другие твердые частицы.
Пиноцитоз — процесс захвата и поглощения капель жидкости с растворенными в ней веществами. Вещества прилипают к белкам мембраны (рецепторам), и капля раствора окружается мембраной, формируя вакуоль. Пиноцитоз и фагоцитоз происходят с затратой энергии АТФ.
- Секреторная. Секреция — выделение клеткой веществ, синтезированных в клетке, во внешнюю среду. Гормоны, полисахариды, белки, жировые капли, заключаются в пузырьки, ограниченные мембраной, и подходят к плазмалемме. Мембраны сливаются, и содержимое пузырька выводится в среду, окружающую клетку.
- Соединение клеток в ткани (за счет складчатых выростов).
- Рецепторная. В мембранах имеется большое число рецепторов — специальных белков, роль которых заключается в передаче сигналов извне внутрь клетки.
Какое строение имеет плазматическая мембрана? Каковы ее функции?
3.9 (77.67%) 43 votesНа этой странице искали :
- строение плазматической мембраны
- каково строение плазматической мембраны
- что такое плазматическая мембрана каково её значение
- плазматическая мембрана строение
- строение и функции плазматической мембраны
Сохрани к себе на стену!
vsesochineniya.ru