Биологические мембраны — Мегаэнциклопедия Кирилла и Мефодия — статья
Биологи́ческие мембра́ны — тонкие (не более 10 нм толщиной) липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который включены молекулы разнообразных белков. Расположены на поверхности клеток (плазматическая мембрана) и внутриклеточных частиц (ядра, митохондрий и др.). В весовом отношении в зависимости от типа мембраны на долю липидов приходится 25-60%, а на долю белков — 40-75%. В состав многих мембран входят углеводы, количество которых может достигать 2-10%.Состав липидов биологических мембран очень разнообразен. Характерными представителями липидов клеточных мембран являются фосфолипиды, сфингомиелины и холестерин (стероидный липид). Характерной особенностью мембранных липидов является разделение их молекулы на две функционально различные части: не полярные, не несущие зарядов хвосты, состоящие из жирных кислот, и заряженные полярные головки. Полярные головки несут на себе отрицательные заряды или могут быть нейтральными. Наличие неполярных хвостов объясняет хорошую растворимость липидов в жирах и органических растворителях. В эксперименте, смешивая с водой выделенные из мембран липиды можно получить бимолекулярные слои или мембраны толщиной около 7, 5 нм, где периферические зоны слоя — это гидрофильные полярные головки, а центральная зона — незаряженные хвосты молекул липидов. Такое же строение имеют все естественные клеточные мембраны. Клеточные мембраны сильно отличаются друг от друга по составу липидов. Например, плазматические мембраны клеток животных богаты холестерином (до 30%), и в них мало лецитина, в то время как мембраны митохондрий богаты фосфолипидами и бедны холестерином. Липидные молекулы могут перемещаться вдоль липидного слоя, могут вращаться вокруг своей оси, а также переходить из слоя в слой. Белки, плавающие в «липидном озере», тоже обладают некоторой латеральной подвижностью. Состав липидов по обе стороны мембраны различен, что определяет асимметричность в строении билипидного слоя.
Мембраны сильно различаются по составу белков, которые собственно и определяют их функциональные свойства. Все мембранные белки можно разделить на три группы: ферменты, рецепторные белки и структурные белки.
В разных мембранах существует характерный набор ферментов. Например, в плазматической мембране локализуется К-Nа-зависимая АТФ-аза, участвующая в транспорте ионов. Рецепторные белки специфически связываются с теми или иными веществами и как бы «узнают» их: это белки-рецепторы для гормонов, для узнавания поверхности соседних клеток, вирусов и т. д. Структурные белки обеспечивают прочность мембраны и связаны с разнообразными белковыми структурами цитоплазмы. Например, в эпителиальных клетках специальные белки плазматической мембраны связываются с элементами цитоскелета и участвуют в образовании целого ряда межклеточных соединений, таких как десмосомы, адгезивные контакты и др.
С мембранами связаны главным образом гликопротеины — молекулы белков, ковалентно связанных с цепочками углеводов. Как правило, молекулы углеводов расположены в наружных слоях мембран. Чаще всего это короткие линейные или разветвленные цепочки, в состав которых входят галактоза, манноза, фруктоза, сахароза, N-ацетилглюкозамин, арабиноза, ксилоза и др.
Строятся все клеточные мембраны (кроме мембран митохондрий и пластид) в гранулярном эндоплазматическом ретикулюме. От него отщепляются мелкие мембранные вакуоли, которые сливаются с мембранами аппарата Гольджи. Мембранные везикулы, продуцируемые, в свою очередь, аппаратом Гольджи, идут на построение плазматической мембраны, секреторных вакуолей и всех других мембранных компонентов клетки. megabook.ruСостав мембран
Основными компонентами мембран являются белки и липиды. На долю углеводов может приходиться около 10% массы мембран, при этом они всегда входят в состав гликолипидов или гликопротеи-нов. Соотношение между белками и липидами в мембранах значительно варьирует — от 20% белка в миелине до 80% в митохондриях. В табл. 1.3 и 1.4 обобщены данные по составу ряда мембран. Плотность мембран прямо пропорциональна содержанию в них белка. Судя по данным изопикнического центрифугирования, чем выше содержание белка в мембране, тем больше ее плотность.
Таблиця 1. Фосфолипидный состав субклеточных мембран печени крысы «
Таблица 2 Белковый и липидиый состав некоторых мембран животных и бактериальных клеток. Л/Б — отношение липка/белок»

Белковый состав мембраны зависит в какой-то степени от метода ее выделения. Некоторые белки непрочно связаны с мембраной и легко удаляются при промывании ее растворами с высокой или, напротив, с низкой ионной силой, щелочными растворами или растворами, содержащими хелатирующие агенты типа ЭДТА. Бывают и случаи, когда трудно сказать, является ли белок мембранным или цитоплазматическим, случайно связавшимся с мембраной в ходе ее выделения.
1. МЕМБРАННЫЕ ЛИПИДЫ
Наиболее поражает в мембранных липидах их огромное разнообразие. Причины этого пока не ясны, хотя становится все более очевидно, что, по-видимому, связано это с тем разнообразием функций, которые липиды выполняют в мембранах. Но, конечно, главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. Основные классы липидов представлены на рис. 1.8; их краткое описание дано ниже.
Глицерофосфолипиды
Это наиболее распространенные липиды. Одна из гидроксильных групп глицерола связана с полярной группировкой, содержащей фосфат, а две другие — с гидрофобными остатками. Номенклатура гли-церидов основана на системе стереоспецифической нумерации. Если глицерол изобразить в проекции Фишера, так что центральная группа будет расположена слева, то атомы углерода будут нумероваться так, как показано на рис. В этом случае в название глицерофосфолипида вводят приставку snдля обозначения положения заместителя. В литературе встречается несколько систем стереохимических обозначений: sn, D/L и R/S. На рис. представлена стереохимия атома С-2 в этих трех системах. Природные фосфолипиды.
У большинства фосфоглицеридов фосфатная группа находится в ял-3-положении глицерола; она обычно связана с какой-либо из групп, представленных на рис.

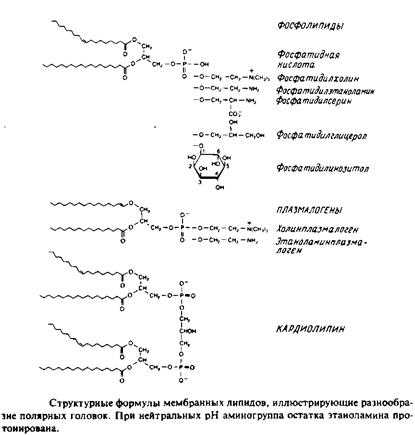
Длинные углеводородные цепи, находящиеся в положениях sn-\ и sn-2, могут присоединяться за счет сложноэфирной и простой эфирной связей. Эти цепи значительно различаются по длине, раз-ветвленности и степени ненасыщенности.
1. 1,2-диацилфосфоглицериды или фосфолипиды. Эти липиды, являющиеся сложными эфирами жирных кислот и глицерола, широко представлены во многих мембранах эукариотических и прокарио-тических клеток, за исключением архебактерий. Фосфатидил-холин является основным компонентом мембран животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы.

Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
Кардиолипины или дифосфатидилглицеролы. По сути это димерные формы фосфолипидов. Они содержатся в боль шом количестве во внутренней мембране митохондрий, в мембране хлоропластов и в некоторых бактериальных мембранах, но редко встречаются в других мембранах.

4. Плазмалогены. Это фосфоглицеролипиды, у которых одна из углеводородных цепей представляет собой простой виниловый эфир. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
Фосфосфинголипиды
Эти липиды имеют такие же полярные головки, как и глицерофосфолипиды, но их гидрофобная часть представлена церамидом. В плазматических мембранах животных клеток широко распространен сфингомиелин. Основными жирнокислотными компонентами в миелине являются кислоты 24:1 и 24:0. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Кроме сфингомиелина известны и другие фосфосфинголипиды, например церамид-1-фосфорилэтаноламин, церамид-1-фосфорилинози-тол и церамид-1-фосфорилглицерол.
Гликоглицеролипиды
Это полярные липиды, у которых в м-З-положении глицерола находится углевод, присоединенный с помощью гликозидной связи, например галактоза. Гликоглицеролипиды широко представлены в мембранах хлоропластов, они обнаружены также в заметных количествах в сине-зеленых водорослях и бактериях. Моногалактозилдиа-цилглицерол был назван «наиболее распространенным в природе полярным липидом», поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов. Для мембран грамположительных бактерий характерны гликоглице-

ролипиды с большим разнообразием Сахаров. Архебактерии также содержат такие липиды, но, как и в случае глицерофосфолипидов, их стереохимическая конфигурация является обращенной, с локализацией гликозидной связи в sn-l-положении глицерола. В мембранах животных клеток гликоглицеролипиды встречаются редко.

Гликосфинголипиды
классифицируют в соответствии с размером углеводной части, которая может быть представлена всего лишь одним моносахаридным остатком, с одной стороны, и очень сложным углеводным полимером — с другой. Моногликозилцерамиды обычно называют цереброзидами. Ганглиозиды представляют собой класс анионных гликосфинголипидов, которые содержат один или несколько остатков сиаловой кислоты, связанных с сахарными остатками церамидолигосахарида. Глобозидами называют нейтральные гликосфинголипиды, которые не содержат остатков отрицательно заряженной сиаловой кислоты.
Гликосфинголипи
mirznanii.com
Химический состав мембран.. Биологическая химия
Химический состав мембран.
Мембраны состоят из липидных и белковых молекул, относительное количество которых у разных мембран широко колеблется. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5%-10% веществ мембраны. Согласно жидкостно-мозаичной модели строения мембраны (Сенджер и Николсон, 1972г.) основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы растворены в липидном бислое и относительно свободно «плавают в липидном море в виде айсбергов на которых растут деревья гликокаликса».
Липиды мембран.
Мембранные липиды – амфифильные молекулы, т.е. в молекуле есть как гидрофильные группы (полярные головки), так и алифатические радикалы (гидрофобные хвосты), самопроизвольно формирующие бислой, в котором хвосты липидов обращены друг к другу. Толщина одного липидного слоя 2,5 нм, из которых 1 нм приходится на головку и 1,5 нм на хвост. В мембранах присутствуют три основных типа липидов: фосфолипиды, гликолипиды и холестерол. Среднее молярное отношение холестерол/фосфолипиды равно 0,3–0,4, но в плазматической мембране это соотношение гораздо выше (0,8–0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липидов и белков.
Фосфолипиды можно разделить на глицерофосфолипиды и сфингофосфолипиды. Наиболее распространенные глицерофосфолипиды мембран – фосфатидилхолины и фосфатидилэтаноламины. Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирнокислотных остатков.
На долю глицерофосфолипидов приходится 2–8% всех фосфолипидов мембран. Наиболее распространенными являются фосфатидилинозитолы.
Специфические фосфолипиды внутренней мембраны митохондрий – кардиолипины (дифосфатидглицеролы), построенные на основе глицерола и двух остатков фосфатидной кислоты, составляют около 22% от всех фосфолипидов митохондриальных мембран.
В миелиновой оболочке нервных клеток в значительных количествах содержатся сфингомиелины.
Гликолипиды мембран представлены цереброзидами и ганглиозидами, в которых гидрофобная часть представлена церамидом. Гидрофильная группа – углеводный остаток – гликозидной связью присоединен к гидроксильной группе первого углеродного атома церамида. В значительных количествах гликолипиды находятся в мебранах клеток мозга, эпителия и эритроцитов. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей и проявляют антигенные свойства.
Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является полярной головкой.
Функции мембранных липидов.
Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других функций. Липиды мембран формируют среду для функционирования мембранных белков, принимающих в ней нативную конформацию.
Некоторые мембранные липиды – предшественники вторичных посредников при передаче гормональных сигналов. Так фосфатидилинозитолдифосфат под действием фосфолипазы С гидролизируется до диацилглицерола и инозитолтрифосфата, являющихся вторичными посредниками гормонов.
Ряд липидов участвует в фиксации заякоренных белков. Примером заякоренного белка является ацетилхолинэстераза, которая фиксируется на постсинаптической мембране к фосфатитилинозитолу.
Поделитесь на страничкеСледующая глава >
bio.wikireading.ru
Биологические мембраны
Состоят из фосфолипидов, гликолипидов, белков и холестерина. В состав липидных компонентов мембран входят только фосфолипиды, а жира и эфиров холестерина в мембранах нет. Липидные участки мембран построены из фосфолипидов (ФЛ), гликолипидов (ГЛ) и ХС.
Мембраны можно рассматривать как белково-липидные комплексы. Белки и липиды, входящие в состав этих комплексов, связаны слабыми типами связей, из которых наиболее часто встречается гидрофобное взаимодействие. Соотношение белков и липидов в различных мембранах разное. Но в большинстве плазматических мембран 50% белков и 50% липидов. Но есть мембраны, в которых много белков: внутренняя мембрана митохондрий на 80% состоит из белков, и только 20% составляют липиды. В миелиновых оболочках нервов, наоборот, 80% — липиды и 20% — белки. ХС встречается в основном в в плазматических мембранах.
В состав мембран входят только ЛИПОИДЫ (сложные липиды): ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и из стероидов — ХОЛЕСТЕРИН (ХС).
Фосфолипиды.
Основу мембран составляют ФОСФОЛИПИДЫ — это липиды, содержащие ФОСФАТНЫЙ ОСТАТОК.
Состоят из четырех компонентов:
1) спирт
2) жирные кислоты
3) фосфат
4) полярная группировка (Если это СЕРИН, то глицерофосфолипид называют ФОСФАТИДИЛСЕРИН, если ХОЛИН, то глицерофосфолипид называют ФОСФАТИДИЛХОЛИН, если ЭТАНОЛАМИН, то глицерофосфолипид называют ФОСФАТИДИЛЭТАНОЛАМИН, если ИНОЗИТ, то глицерофосфолипид называют ФОСФАТИДИЛИНОЗИТ).
ОБЩАЯ ФОРМУЛА ГЛИЦЕРОФОСФОЛИПИДОВ :
В состав фосфолипидов могут входить 2 спирта: глицерин (глицерофосфолипиды) и сфингозин (сфингофосфолипиды, сфингомиелины). Все компоненты соединены эфирными связями. Кроме разделения на основе содержания той или иной полярной группы, их делят на основе содержащегося в них спирта:
1. ГЛИЦЕРОФОСФОЛИПИДЫ (ГФЛ) — содержат спирт глицерин.
Все они относятся к L-ряду. Есть асимметрический углеродный атом (на рисунке обозначен звездочкой). Полярная группировка может быть представлена аминокислотой серином (фосфатидилсерин), холином (фосфатидилхолин, другое название – лецитин), этаноламином (фосфатидилэтаноламин), инозитолом (фосфатидилинозитол), глицерином (полиглицерофосфатиды).
В природных фосфолипидах R1 и R2 — разные. R1 — насыщенная жирная кислота, R2.- ненасыщенная жирная кислота. Однако, есть и исключения: основным липидным компонентом легочного сурфактанта является ГФЛ, у которого и R1, и R2 – радикалы пальмитиновой кислоты, а полярная группировка – холин.
2. СФИНГОФОСФОЛИПИДЫ (СФЛ) — содержат спирт сфингозин: СФИНГОМИЕЛИНЫ.
Сфингофосфолипиды бывают различными по строению, но имеют общие черты. Молекула сфингофосфолипида содержит сфингозин, жирную кислоту, фосфорную кислоту и полярную группировку.
ОБЩАЯ ФОРМУЛА СФЛ представлена на рисунке.
Сфингозин — это 2-хатомный непредельный аминоспирт.
Жирная кислота присоединена пептидной связью к аминогруппе сфингозина.
Фосфолипиды — это амфифильные вещества. Расположение гидрофильных и гидрофобных участков особое. Гидрофильные участки (остаток фосфорной кислоты и полярная группировка) образуют «головку», а гидрофобные радикалы жирных кислот (R1 и R2) образуют «хвосты».
Поэтому молекулу фосфолипида обозначают:
ГЛИКОЛИПИДЫ.
Состоят из сфингозина, жирной кислоты и молекулы какого-либо углевода. Если в формулу СФЛ вместо фосфорной кислоты поставить какой-нибудь углевод, то получим формулу ГЛ. Гликолипиды тоже имеют гидрофильную «головку» и 2 гидрофобных «хвоста». Общая схема их строения представлена на рисунке:
Гликолипиды классифицируют в зависимости от строения углеводного компонента.
Различают 2 группы гликолипидов:
1. ЦЕРЕБРОЗИДЫ. В качестве углеводного компонента содержат какой-либо моносахарид (глюкоза, галактоза), либо дисахарид, или нейтральный небольшой олигосахарид.
2. ГАНГЛИОЗИДЫ. Углеводным компонентом является олигосахарид, состоящий из разных мономеров, как самих моносахаридов, так и их производных. Этот олигосахарид обязательно кислый, в его состав обязательно входит сиаловая кислота. Благодаря определенной последовательности мономеров, олигосахариды в составе ганглиозида придают молекуле выраженные антигенные свойства.
СТЕРОИДЫ.
Делятся на 2 группы.
1. Стерины (в их составе полициклическая стуктура стерана).
2. Стериды (эфиры холестерина и высших жирных кислот).
Свойства стероидов.
Стерины содержат гидроксильную группу (-ОН), поэтому они немножко гидрофильны, но всётаки их молекулы в основном гидрофобны. К ним относится холестерин.
Холестерин является полициклическим веществом. Преобладают гидрофобные свойства, но есть одна ОН-группа.
Стериды являются полностью гидрофобными веществами.
ФЛ и ГЛ вместе называют «полярные липиды». Если смешать полярные липиды с водой, то наблюдается взаимодействие между ними и при определенных условиях полярные липиды могут спонтанно образовывать бимолекулярный слой (бислой), схематично представленный на рисунке:
Между «головками» ионные, водородные связи, между «хвостами» — гидрофобное взаимодействие. Липидная часть мембраны состоит из таких липидов.Свойства бислоя липидов:
1. Маленькая толщина — в 2 молекулы (4-13 нм)
2. Высокая эластичность. При 37оС липоиды находятся в жидком состоянии. Значит, возможны перемещения, однако скорость их диффузии в 100 раз меньше, чем у молекул воды.
Виды движений: а)в пределах своего монослоя; б) вращательные движения; в) флуктуация «хвостов».
Переход молекулы из одного слоя в другой — это редкое явление.
В настоящее время мембрана рассматривается как жидкокристаллическая структура. Наряду с диффузией имеется упорядоченность.
3. Третье свойство бислоя: низкая электропроводность. Поэтому липидный бислой является хорошим диэлектриком.
4. Четвертое свойство связано с избирательной проницаемостью липидного бислоя.
Сквозь него могут свободно проходить небольшие электронейтральные молекулы кислорода, углекислоты, азота, а также вещества, имеющие гидрофобную природу. Например, стероидные гормоны, обладающие внутриклеточным механизмом действия, широко применяются в медицине, в том числе и местно — они способны проникать даже через кожу, слизистую оболочку глаз (лечение кожных и глазных заболеваний). Органические растворители тоже проникают внутрь через кожу или легкие при вдыхании паров. Поэтому возможны отравления этими веществами через кожу, слизистые оболочки, дыхательные пути.
Заряженные молекулы через билипидный слой не проникают. Поэтому транспорт таких молекул осуществляют специальные мембранные транспортные белки.
Кроме липоидов, мембрана содержит и белки.
Встречаются 2 типа белков:
1. Периферические белки — взаимодействуют с «головками» полярных липидов электростатически.
2. Интегральные белки — взаимодействуют как с «головками» липидов, так и с гидрофобными «хвостами». В интегральных белках преобладают гидрофобные аминокислоты.
Белки, как и липоиды, слабо связаны с мембранами. Поэтому периферические белки часто сравнивают с льдинами, которые плавают по морю, а интегральные — с айсбергами. Имеются также специальные белки («якорные»), которые прикрепляют мембрану к белкам цитоскелета.
studfiles.net
Биологические мембраны
Состоят из фосфолипидов, гликолипидов, белков и холестерина. В состав липидных компонентов мембран входят только фосфолипиды, а жира и эфиров холестерина в мембранах нет. Липидные участки мембран построены из фосфолипидов (ФЛ), гликолипидов (ГЛ) и ХС.
Мембраны можно рассматривать как белково-липидные комплексы. Белки и липиды, входящие в состав этих комплексов, связаны слабыми типами связей, из которых наиболее часто встречается гидрофобное взаимодействие. Соотношение белков и липидов в различных мембранах разное. Но в большинстве плазматических мембран 50% белков и 50% липидов. Но есть мембраны, в которых много белков: внутренняя мембрана митохондрий на 80% состоит из белков, и только 20% составляют липиды. В миелиновых оболочках нервов, наоборот, 80% — липиды и 20% — белки. ХС встречается в основном в в плазматических мембранах.
В состав мембран входят только ЛИПОИДЫ (сложные липиды): ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и из стероидов — ХОЛЕСТЕРИН (ХС).
Фосфолипиды.
Основу мембран составляют ФОСФОЛИПИДЫ — это липиды, содержащие ФОСФАТНЫЙ ОСТАТОК.
Состоят из четырех компонентов:
1) спирт
2) жирные кислоты
3) фосфат
4) полярная группировка (Если это СЕРИН, то глицерофосфолипид называют ФОСФАТИДИЛСЕРИН, если ХОЛИН, то глицерофосфолипид называют ФОСФАТИДИЛХОЛИН, если ЭТАНОЛАМИН, то глицерофосфолипид называют ФОСФАТИДИЛЭТАНОЛАМИН, если ИНОЗИТ, то глицерофосфолипид называют ФОСФАТИДИЛИНОЗИТ).
ОБЩАЯ ФОРМУЛА ГЛИЦЕРОФОСФОЛИПИДОВ:
В состав фосфолипидов могут входить 2 спирта: глицерин (глицерофосфолипиды) и сфингозин (сфингофосфолипиды, сфингомиелины). Все компоненты соединены эфирными связями. Кроме разделения на основе содержания той или иной полярной группы, их делят на основе содержащегося в них спирта:
1. ГЛИЦЕРОФОСФОЛИПИДЫ (ГФЛ) — содержат спирт глицерин.
Все они относятся к L-ряду. Есть асимметрический углеродный атом (на рисунке обозначен звездочкой). Полярная группировка может быть представлена аминокислотой серином (фосфатидилсерин), холином (фосфатидилхолин, другое название – лецитин), этаноламином (фосфатидилэтаноламин), инозитолом (фосфатидилинозитол), глицерином (полиглицерофосфатиды).
В природных фосфолипидах R1 и R2 — разные. R1 — насыщенная жирная кислота, R2.- ненасыщенная жирная кислота. Однако, есть и исключения: основным липидным компонентом легочного сурфактанта является ГФЛ, у которого и R1, и R2 – радикалы пальмитиновой кислоты, а полярная группировка – холин.
2. СФИНГОФОСФОЛИПИДЫ (СФЛ) — содержат спирт сфингозин: СФИНГОМИЕЛИНЫ.
Сфингофосфолипиды бывают различными по строению, но имеют общие черты. Молекула сфингофосфолипида содержит сфингозин, жирную кислоту, фосфорную кислоту и полярную группировку.
ОБЩАЯ ФОРМУЛА СФЛ представлена на рисунке.
Сфингозин — это 2-хатомный непредельный аминоспирт.
Жирная кислота присоединена пептидной связью к аминогруппе сфингозина.
Фосфолипиды — это амфифильные вещества. Расположение гидрофильных и гидрофобных участков особое. Гидрофильные участки (остаток фосфорной кислоты и полярная группировка) образуют «головку», а гидрофобные радикалы жирных кислот (R1 и R2) образуют «хвосты».
Поэтому молекулу фосфолипида обозначают:
ГЛИКОЛИПИДЫ.
Состоят из сфингозина, жирной кислоты и молекулы какого-либо углевода. Если в формулу СФЛ вместо фосфорной кислоты поставить какой-нибудь углевод, то получим формулу ГЛ. Гликолипиды тоже имеют гидрофильную «головку» и 2 гидрофобных «хвоста». Общая схема их строения представлена на рисунке:
Гликолипиды классифицируют в зависимости от строения углеводного компонента.
Различают 2 группы гликолипидов:
1. ЦЕРЕБРОЗИДЫ. В качестве углеводного компонента содержат какой-либо моносахарид (глюкоза, галактоза), либо дисахарид, или нейтральный небольшой олигосахарид.
2. ГАНГЛИОЗИДЫ. Углеводным компонентом является олигосахарид, состоящий из разных мономеров, как самих моносахаридов, так и их производных. Этот олигосахарид обязательно кислый, в его состав обязательно входит сиаловая кислота. Благодаря определенной последовательности мономеров, олигосахариды в составе ганглиозида придают молекуле выраженные антигенные свойства.
СТЕРОИДЫ.
Делятся на 2 группы.
1. Стерины (в их составе полициклическая стуктура стерана).
2. Стериды (эфиры холестерина и высших жирных кислот).
Свойства стероидов.
Стерины содержат гидроксильную группу (-ОН), поэтому они немножко гидрофильны, но всётаки их молекулы в основном гидрофобны. К ним относится холестерин.
Холестерин является полициклическим веществом. Преобладают гидрофобные свойства, но есть одна ОН-группа.
Стериды являются полностью гидрофобными веществами.
ФЛ и ГЛ вместе называют «полярные липиды». Если смешать полярные липиды с водой, то наблюдается взаимодействие между ними и при определенных условиях полярные липиды могут спонтанно образовывать бимолекулярный слой (бислой), схематично представленный на рисунке:
Между «головками» ионные, водородные связи, между «хвостами» — гидрофобное взаимодействие. Липидная часть мембраны состоит из таких липидов.Свойства бислоя липидов:
1. Маленькая толщина — в 2 молекулы (4-13 нм)
2. Высокая эластичность. При 37оС липоиды находятся в жидком состоянии. Значит, возможны перемещения, однако скорость их диффузии в 100 раз меньше, чем у молекул воды.
Виды движений: а)в пределах своего монослоя; б) вращательные движения; в) флуктуация «хвостов».
Переход молекулы из одного слоя в другой — это редкое явление.
В настоящее время мембрана рассматривается как жидкокристаллическая структура. Наряду с диффузией имеется упорядоченность.
3. Третье свойство бислоя: низкая электропроводность. Поэтому липидный бислой является хорошим диэлектриком.
4. Четвертое свойство связано с избирательной проницаемостью липидного бислоя.
Сквозь него могут свободно проходить небольшие электронейтральные молекулы кислорода, углекислоты, азота, а также вещества, имеющие гидрофобную природу. Например, стероидные гормоны, обладающие внутриклеточным механизмом действия, широко применяются в медицине, в том числе и местно — они способны проникать даже через кожу, слизистую оболочку глаз (лечение кожных и глазных заболеваний). Органические растворители тоже проникают внутрь через кожу или легкие при вдыхании паров. Поэтому возможны отравления этими веществами через кожу, слизистые оболочки, дыхательные пути.
Заряженные молекулы через билипидный слой не проникают. Поэтому транспорт таких молекул осуществляют специальные мембранные транспортные белки.
Кроме липоидов, мембрана содержит и белки.
Встречаются 2 типа белков:
1. Периферические белки — взаимодействуют с «головками» полярных липидов электростатически.
2. Интегральные белки — взаимодействуют как с «головками» липидов, так и с гидрофобными «хвостами». В интегральных белках преобладают гидрофобные аминокислоты.
Белки, как и липоиды, слабо связаны с мембранами. Поэтому периферические белки часто сравнивают с льдинами, которые плавают по морю, а интегральные — с айсбергами. Имеются также специальные белки («якорные»), которые прикрепляют мембрану к белкам цитоскелета.
studfiles.net
Мембраны, их состав, структура. Мембранный транспорт. — КиберПедия
Биологическими мембранами называют функциональные структуры клеток, ограничивающих цитоплазму и большинство внутриклеточных структур. Мембраны также образуют внутри клетки единую систему канальцев и полостей. Толщина биологических мембран составляет 7-10 нм, однако вследствие большой общей площади мембран и их высокой плотности, их масса составляет более 50 % сухой массы клетки. Биологические мембраны являются высокоизбирательными барьерами проницаемости. Поток молекул в клетку и из клетки, в органоид и из нее, контролируется находящимися в мембранах специфическими системами транспорта. Транспортные процессы в мембране регулируют объем клетки и поддерживают ионный состав, необходимый для работы ферментативныхсистем, т.е. гомеостаз клетки. Транспортные процессы также создают ионные градиенты, необходимые для создания мембранного потенциала, поддержания возбудимости, а также транспорта некоторых молекул и ионов. Системы транспорта в мембране переносят из внешней среды и концентрируют в компартментах клетки вещества, необходимые для функционирования клетки. Так как клетки и внутриклеточные структуры, с точки зрения термодинамики, открытые системы, они постоянно обмениваются с окружающей средой и веществом и энергией. Поэтому транспорт вещества и энергии через мембраны — необходимое условие существования живых систем.
Состав и структура биомембран.
Состав мембран зависит от их типа и функций, однако во всех типах биологических мембран основными структурными компонентами являются молекулы липидов и белков. Некоторые мембраны содержат также и углеводы, которые связаны или с белками (гликопротеины) или с липидами (гликолипиды). Важным структурным компонентом мембран является вода. Взаимодействие молекул липидов, белков с водой определяет специфические структурно-функциональные свойства мембраны и определяет стабильность мембранных структур.
В состав биологических мембран входят молекулы, относящиеся к различным классам липидов, а также стероиды. Мембранные липиды имеют сравнительно небольшую полярную (заряженную) головку и длинные неполярные (незаряженные) углеводородные цепи. Жирные кислоты, входящие в состав липидов мембран, могут быть насыщенными и ненасыщенными двойными связями и обычно имеют 14-22 углеродных атома.
У фосфолипидов (фосфоглицеридов) неполярная часть представлена двумя остатками жирных кислот, этерефицирующих две гидроксильные группы глицерола. Третья гидроксильная группа образует полярную головку – остаток фосфорной кислоты. К остатку фосфорной кислоты может быть присоединен остаток аминоспирта (например, фосфатидилхолин), сахара (фосфатидилинозитол), аминокислоты (фосфатидилсерин).
Сфинголипиды построены из одного остатка жирной кислоты, из одного остатка длинноцепочечного аминоспирта сфингозина (или его производного). Полярная головка этих молекул представлена остатком спирта.
Из стероидов наиболее распространенным компонентом мембран является холестерин (холестерол) и его производные. Содержание холестерина характерно для мембран эукариотических клеток, у большинства прокариот он не обнаружен.
Белковый состав мембран также исключительно многообразен. Мембраны многих клеток содержат большое число белков, с молекулярной массой 10 до 250 кДа. Молекулы белков могут быть либо частично, либо целиком, погружены в липидный бислой или могут пронизывать его насквозь. Это так называемые интегральные белки. Белки, которые слабо удерживаются на мембране за счет слабых, в основном электростатических, взаимодействий, называются периферическими. Молекулы белков в составе мембран выполняют ферментативные, транспортные, регуляторные и опорно-строительную функции. Относительное содержание белков и липидов в составе мембран зависит от типа клеток и от специфики мембранных функций. Так, в составе внутриклеточных мембран (митохондрий, микросом) содержание белка выше, чем содержание липидов. В мембранах нервных клеток млекопитающих содержание белка составляет всего 20-25 % по сухой массе ( табл.1)
Таблица 1
Относительное содержание белков и липидов в составе различных мембран (% от сухой массы)
| Вид мембран | белки | липиды |
| Миелиновая оболочка нервных клеток млекопитающих | ||
| Плазмалемма эритроцитов человека | ||
| Плазмалемма клеток печени млекопитающих | ||
| Наружная мембрана митохондрий млекопитающих | ||
| Внутренняя мембрана митохондрий млекопитающих | ||
| Микросомы клеток печени млекопитающих |
Относительное содержание представителей того иного класса липидов, белковых молекул, также зависит от структурных особенностей и функциональных свойств мембраны. Так, мембрана нервных клеток, содержит небольшое количество белка ( 18 % массы мембраны). В плазматической мембране большинства клеток содержание белка достигает до 50 % массы, так как различные ионные насосы, каналы, рецепторы, ферменты представлены, в основном, молекулами белков. Наиболее высоким содержанием белков характеризуются мембраны внутренние митохондрий и хлоропластов, где располагаются электронно- транспортные цепи и происходит синтез АТФ. В составе мембран большинства типов клеток наиболее распространены следующие молекулы липидов: фосфатидилхолин (лецитин), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол. Мембраны эукариотических клеток богаты также холестерином.
В таблице 2 приведены данные о липидном составе различных мембран клеток млекопитающих.
Таблица 2
Содержание липидов в составе плазматической мембраны и мембран органоидов животной клетки ( в % к общей массе липидов)
| Липиды | Плазмалемма | Митохондрии | Лизосомы | Ядро | ЭПС | Аппа рат Гольджи |
| Фосфатидил холин | ||||||
| Сфингомиелин | ||||||
| Фосфатидилэтаноламин | 11,5 | 28,5 | 12,5 | 16.5 | ||
| Фосфатидилсерин | 3,5 | 2,5 | ||||
| Фосфатидилинозитол | 2,5 | 7,5 | ||||
| Лизофосфатидилхолин | 2,5 | 1,5 | ||||
| Дифосфатидилглицерин | ||||||
| Холестерин и ее эфиры | ||||||
| Свободные жирные кислоты | - | - | ||||
| Другие липиды | 2,5 | 5,5 |
Воду, входящую в состав мембран, подразделяют на связанную, захваченную и свободную. Связанная вода — это вода гидратных оболочек ионов и полярных участков липидов и белков. Гидратные оболочки основных структурообразующих липидов состоят из 10-12 молекул воды. Эта вода осмотически и химически неактивна и не способна растворять какие — либо вещества. Захваченная вода находится в основном между двумя слоями в липидов мембраны. По подвижности, химической активности она занимает промежуточное положение между связанной и свободной водой. Свободная вода входит в состав мембраны в виде самостоятельной фазы и обладает всеми свойствами жидкой воды.
cyberpedia.su
Клеточная мембрана состав — Справочник химика 21
Все биологические мембраны, включая плазматическую мембран и внутренние мембраны эукариотических клеток, имеют общие структурные особенности они представляют собой ансамбли липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий. Благодаря этим взаимодействиям поддерживается структурная целостность мембран Однако сами по себе клеточные мембраны являются подвижными, текучими структурами и большинство входящих в их состав молекул способны перемещаться в плоскости мембраны. Как показано на рис. 6-1, липидные молекулы образуют непрерывный двойной слой толщиной около 5 нм. Липидный бислой — это основная структура мембраны, которая и создает относительно непроницаемый барьер для большинства водорастворимых молекул. Белковые молекулы как бы растворены в липидном бислое. С их помощью выполняются разнообразные функции мембраны. Одни мембранные белки обеспечивают транспорт молекул внутрь клетки или из нее, другие являются ферментами и катализируют ассоциированные с мембраной реакции. Еще один класс белков осуществляет структурную связь плазматической мембраны с цитоскелетом, с одной стороны, и(или) с внеклеточным матриксом либо с соседней клеткой — с другой. Отдельную группу составляют белки, выполняющие роль рецепторов для получения и преобразования химических сигналов из окружающей среды. Как и следовало ожидать, мембраны асимметричны оба их слоя различаются по липидному и белковому составу, что отражает, по-видимому, функциональные различия их поверхностей. [c.349]В состав клеточных мембран входят в основном белки и липиды, среди- которых преобладают фосфолипиды, составляющие 40—90 % от общего количества липидов в мембране. Строение биомембраны интенсивно изучается в настоящее время. В одной из моделей клеточная мембрана рассматривается как липидный бислой. В таком бислое углеводородные хвосты липидов за счет гидрофобных взаимодействий удерживаются друг возле друга в вытянутом состоянии во внутренней полости, образуя двойной углеводородный слой. Полярные группы липидов располагаются на внешней поверхности бислоя (рис. 14.2). [c.466]
Биологические функции углеводов разнообразны. Углеводы служат источником энергии для клетки, так как при их окислении выделяется много теплоты. Углеводы входят в состав веществ, образующих клеточные мембраны, им принадлежит ис- [c.43]
Большинство прокариот имеет ригидную клеточную стенку, под которой расположена цитоплазматическая мембрана. Состав и строение клеточной стенки — важный систематический признак, по которому прокариоты подразделяют на следующие группы грам-положительные, грамотрицательные и не имеющие клеточной стенки. Своеобразным строением и составом клеточной стенки характеризуются археи. Грамположительные бактерии отличаются от храмотрицательных большим (до 40 раз) содержанием муреина (пептидогликана) в клеточной стенке и отсутствием внешней мем- браны. Археи муреина не синтезируют, но некоторые образуют псевдомуреин. [c.7]
Состав внеклеточной жидкости близок к составу морской воды в пред-кембрийскую эпоху, когда появились животные с замкнутой системой кровообращения. С тех пор соленость моря продолжала возрастать, тогда как состав внеклеточной жидкости остался постоянным. Основным катионом во внеклеточной жидкости является ион Ка , а из анионов преобладают СГ и НСОВнутри клеток преобладают катион и анион НРО Для соблюдения физико-химического закона электронейтральности, которому подчиняется любой живой организм в целом, некоторый недостаток неорганических анионов компенсируется анионами органических кислот (молочной, лимонной и др.) и кислых белков, несущих отрицательный заряд при физиологических значениях pH. Если вне клетки органические анионы компенсируют незначительную нехватку отрицательного заряда, то внутри клетки они должны компенсировать около 25 % положительных зарядов, создаваемых неорганическими катионами. Поскольку клеточные мембраны легко проницаемы для воды, то они могут разрушаться при незначительных различиях в давлении жидкости внутри и снаружи клеточной мембраны. Поэтому осмотическое давление внутри клетки должно быть равно таковому во внеклеточной жидкости, т. е. живая клетка подчиняется закону изоосмоляльности. Повышенное содержание катионов по отношению к концентрации анионов во внеклеточных жидкостях в сравнении с внутриклеточными средами приводит к тому, что наружная поверхность мембран клеток оказывается заряжена положительно относительно ее внутренней поверхности, и это имеет огромное биологическое значение (см. главу 15). В биологических жидкостях концентрацию осмотически активных частиц (независимо от их заряда, размера и массы) выражают в единицах осмоляльности — миллиосмомолях на 1 кг воды. Так как главные катионы и анионы внутриклеточных жидкостей многозарядные, то (при одинаковых осмоляльностях) концентрация электролитов, выраженная в миллиэквивалентах на 1 л, будет значительно выше внутри клетки, чем во внеклеточных жидкостях, где в основном содержатся однозарядные ионы. [c.180]
Функционирует А. в составе сложного комплекса, состоящего из гормонального рецептора (К), расположенного на внеш. стороне клеточной мембраны, внутримембранного регуляторного белка (С), в состав к-рого входит остаток [c.31]
Эта система участвует не только в синтезе ферментов, которые сек-ретируются клеткой, но и в образовании новых мембран. По-видимому, шероховатый ЭР поставляет мембранный материал гладкому ЭР и аппарату Гольджи, а компоненты мембран Гольджи включаются в состав наружной клеточной мембраны. В растительных клетках наружные мембраны митохондрий и мембраны, окружающие вакуоли, также образуются непосредственно из ЭР [19]. Компоненты наружных клеточных мембран, вероятно, могут использоваться повторно, включаясь в соответствующую структуру в ходе эндоцитоза [20]. [c.33]
Мембранные рецепторы выполняют функции узнавания (иммунокомпе-тентная система), адгезии (обеспечение межклеточных контактов, формирование тканей), регуляции активности ионных каналов (электрическая возбудимость, создание мембранного потенциала). Мембранные ферменты в составе бислоя приобретают большую стабильность и способность к осуществлению реакций, которые в гидрофильном окружении протекали бы с весьма малой скоростью. Липидное окружение предоставляет таким белкам привилегированные условия функционирования, но и накладывает ограничения на поведение белковых ассоциатов последнее сильно зав
www.chem21.info