Какую функцию выполняет клеточная мембрана
Клеточная мембрана — молекулярная структура, которая состоит из липидов и белков. Главные её свойства и функции:
- отделение содержимого любой клетки от внешней среды, гарантируя её целостность;
- управление и налаживание обменом между средой и клеткой;
- внутриклеточные мембраны разбивают клетку на специальные отсеки: органеллы или компартменты.
Слово «мембрана» на латыни означает «пленка». Если говорить о клеточной мембране, то это совокупность двух пленок, которые обладают различными свойствами.
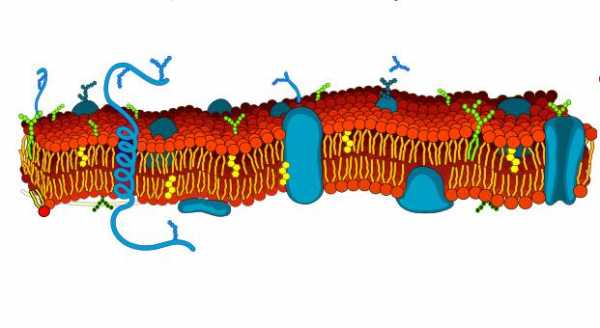
Биологическая мембрана включает в себя три вида белков:
- Периферические – расположены на поверхности пленки;
- Интегральные – целиком пронизывают мембрану;
- Полуинтегральные – одним концом проникают внутрь билипидного слоя.
Какие функции выполняет клеточная мембрана
1. Клеточная стенка — прочная оболочка клетки, которая находится снаружи от цитоплазматической мембраны. Она выполняет защитные, транспортные и структурные функции. Присутствует у многих растений, бактерий, грибов и архей.
2. Обеспечивает барьерную функцию, то есть избирательный, регулируемый, активный и пассивный обмен веществ с внешней средой.
3. Способна передавать и сохранять информации, а также принимает участие в процессе размножения.
4. Выполняет транспортную функцию, которая может через мембрану транспортировать вещества в клетку и из клетки.
5. Клеточная мембрана имеет одностороннюю проводимость. Благодаря этому, молекулы воды могут без задержек проходить через клеточную мембрану, а молекулы прочих веществ проникают выборочно.
6. С помощью клеточной мембраны происходит получение воды, кислорода и питательных веществ, а через неё удаляются продукты клеточного обмена.
7. Выполняет клеточный обмен через мембраны, и может исполнять их с помощью 3 главных типов реакций: пиноцитоз, фагоцитоз, экзоцитоз.
8. Мембрана обеспечивает специфику межклеточных контактов.
9. В мембране присутствуют многочисленные рецепторы, которые способны воспринимать химические сигналы — медиаторы, гормоны и множество других биологических активных веществ. Так она в силах изменить метаболическую активность клетки.
10. Основные свойства и функции клеточной мембраны:
- Матричная
- Барьерная
- Транспортная
- Энергетическая
- Механическая
- Ферментативная
- Рецепторная
- Защитная
- Маркировочная
- Биопотенциальная
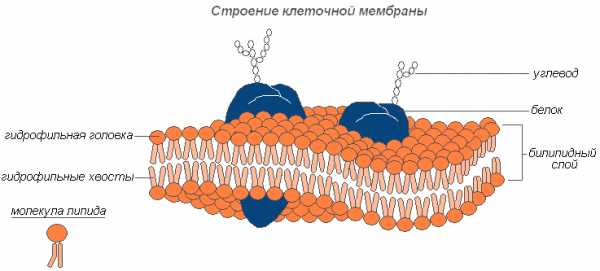
Какую функцию выполняет в клетке плазматическая мембрана?
- Отграничивает содержимое клетки;
- Осуществляет поступление веществ в клетку;
- Обеспечивает удаление ряда веществ из клетки.
Структура мембраны клетки
Клеточные мембраны включают липиды 3 классов:
- Гликолипиды;
- Фосфолипиды;
- Холестерол.
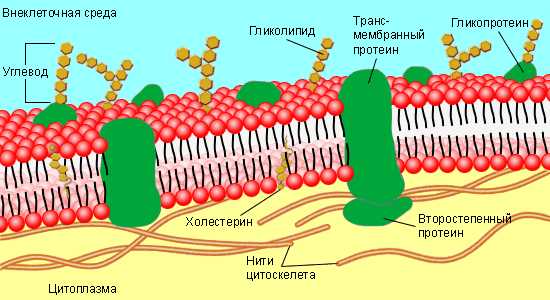
В основном мембрана клетки состоит из белков и липидов, и имеет толщину не более 11 нм. От 40 до 90% всех липидов составляют фосфолипиды. Также важно отметить гликолипиды, которые являются одним из основных компонентов мембраны.
Структура клеточной мембраны трехслойна. В центре располагается однородный жидкий билипидный слой, а белки закрывают его с двух сторон (как мозаику), отчасти проникая в толщу. Также белки необходимы для мембраны, чтобы пропускать внутрь клеток и транспортировать из них наружу особые вещества, которые не могут проникнуть через жировой слой. Например, ионы натрия и калия.
Строение клетки — видео
pristor.ru
Клеточная мембрана, ее основные функции. Чудо творения.
Функции клеточной или плазматической мембраны
text_fields
text_fields
arrow_upward
Клетки отделены от внутренней среды организма клеточной или плазматической мембраной.
Мембрана обеспечивает:
1) Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток;
2) Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала;
3) Специфику межклеточных контактов.
Благодаря наличию в мембране многочисленных рецепторов, воспринимающих химические сигналы — гормоны, медиаторы и другие биологически активные вещества, она способна изменять метаболическую активность клетки. Мембраны обеспечивают специфику иммунных проявлений, благодаря наличию на них антигенов — структур, вызывающих образование антител, способных специфически связываться с этими антигенами.
Структура мембраны клетки
text_fields
text_fields
arrow_upward
Мембрана клетки — эластичная структура, толщиной от 7 до 11 нм (рис.1.1). Она состоит, в основном, из липидоа и белков. От 40 до 90% всех липидов составляют фосфолипиды — фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, сфингомиелин и фосфатидилинозит. Важным компонентом мембраны являются гликолипиды, представленные цереброзидами, сульфатидами, ганглиозидами и холестерином.
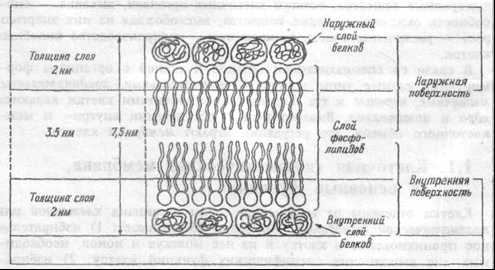 Рис. 1.1 Организация мембраны.
Рис. 1.1 Организация мембраны.Основной структурой мембраны клетки является двойной слой фосфолипидных молекул. За счет гидрофобных взаимодействий углеводные цепочки липидных молекул удерживаются друг возле друга в вытянутом состоянии. Группы же фосфолипидных молекул обоих слоев взаимо действуют с белковыми молекулами, погруженными в липидную мембрану. Благодаря тому, что большинство липидных компонентов бислоя находится в жидком состоянии, мембрана обладает подвижностью, совершает волнообразные движения. Ее участки, а также белки, погруженные в липидный бислой, перемешаются из одной ее части в другую. Подвижность (текучесть) мембран клеток облегчает процессы транспорта веществ через мембрану.
Белки мембраны клеток представлены, в основном, гликопротеинами. Различают:
• интегральные белки
• периферические белки, прикрепленные только к поверхности мембраны, в основном, к внутренней ее части.
Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная фосфатазы и др.). Но некоторые энзимы также представлены интегральными белками — АТФ-аза.
Интегральные белки обеспечивают селективный обмен ионов через каналы мембран между экстрацеллюлярной и интрацеллюлярной жидкостью, а также действуют как белки — переносчики крупных молекул.
Рецепторы и антигены мембраны могут быть представлены как интегральными, так и периферическими белками.
Белки, примыкающие к мембране с цитоплазматической стороны, относятся к цитоскелету клетки. Они могут прикрепляться к мембранным белкам.
Так, белок полосы 3 (номер полосы при электрофорезе белков) эритроцитарных мембран объединяется в ансамбль с другими молекулами цитоскелета — спектрином через низкомолекулярный белок анкирин (рис. 1.2).
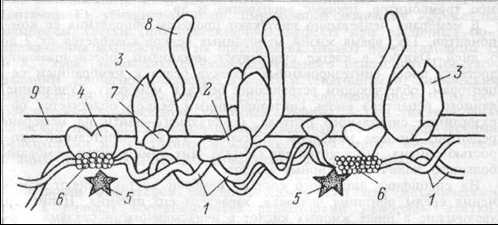 Рис. 1.2 Схема расположения белков в примембранном цитоскелете эритроцитов.
Рис. 1.2 Схема расположения белков в примембранном цитоскелете эритроцитов.1 — спектрин; 2 — анкирин; 3 — белок полосы 3; 4 — белок полосы 4,1; 5 — белок полосы 4,9; 6 — олигомер актина; 7 — белок 6; 8 — гпикофорин А; 9 — мембрана.
Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется актин.
Актин образует микрофиламенты, представляющие собой сократительный аппарат цитоскелета.
Цитоскелет позволяет клетке проявлять гибкоэластические свойства, обеспечивает дополнительную прочность мембраны.
Большинство интегральных белков — гликопротеины. Их углеводная часть выступает из клеточной мембраны наружу. Многие гликопротеины обладают большим отрицательным зарядом из-за значительного содержания сиаловой кислоты (например, молекула гликофорина). Это обеспечивает поверхности большинства клеток отрицательный заряд, способствуя отталкиванию других отрицательно заряженных объектов. Углеводные выступы гликопротеинов являются носителями антигенов групп крови, других антигенных детерминант клетки, они действуют как рецепторы, связывающие гормоны. Гликопротеины образуют адгезивные молекулы, обуславливающие прикрепление клеток одна к другой, т.е. тесные межклеточные контакты.
Особенности обмена веществ в мембране
text_fields
text_fields
arrow_upward
Мембранные компоненты подвержены многим метаболическим превращениям под влиянием ферментов, расположенных на их мембране или внутри ее. К ним относятся окислительные ферменты, играющие важную роль в модификации гидрофобных элементов мембран — холестерина и др. В мембранах же при активации ферментов — фосфолипаз происходит образование из арахидоновой кислоты биологически активных соединений — простагландинов и их производных. В результате активации метаболизма фосфолипидов в мембране образуются тромбоксаны, лейкотриены, оказывающие мощное воздействие на адгезию тромбоцитов, процесс воспаления и др.
В мембране непрерывно протекают процессы обновления ее компонентов. Так, время жизни мембранных белков колеблется от 2 до 5 дней. Однако в клетке существуют механизмы, обеспечивающие доставку вновь синтезированных молекул белка к мембранным рецепторам, облегчающим встраивание белка в мембрану. «Узнавание» данного рецептора вновь синтезированным белком облегчается образованием сигнального пептида, помогающего найти на мембране рецептор.
Липиды мембраны отличаются также значительной скоростью обмена, что требует для синтеза этих компонентов мембраны большого количества жирных кислот.
На специфику липидного состава мембран клеток влияют изменения среды обитания человека, характера его питания.
Например, увеличение в пище жирных кислот с ненасыщенными связями увеличивает жидкое состояние липидов мембран клеток различных тканей, приводит к благоприятному для функции мембраны клетки изменению отношения фосфолипидов к сфингомиелинам и липидов к белкам.
Избыток холестерина в мембранах, напротив, увеличивает микровязкость их бислоя фосфолипидных молекул, понижая скорость диффузии некоторых веществ через мембраны клеток.
Пища, обогащенная витаминами А, Е, С, Р улучшает обмен липидов в мембранах эритроцитов, снижает микровязкость мембран. Это повышает деформируемость эритроцитов, облегчает выполнение ими транспортной функции (глава 6).
Дефицит жирных кислот и холестерина в пище нарушает липидный состав и функции мембран клеток.
Например, дефицит жиров нарушает функции мембраны нейтрофилов, что угнетает их способность к движению и фагоцитозу (активный захват и поглощение микроскопических инородных живых объектов и твердых частиц одноклеточными организмами или некоторыми клетками).
В регулировании липидного состава мембран и их проницаемости, регуляции пролиферации клеток важную роль играют активные формы кислорода, образующиеся в клетке сопряженно с нормально протекающими метаболическими реакциями (микросомальным окислением и др.).
Образующиеся активные формы кислорода — супероксидный радикал (О2), перекись водорода (H2О2) и др. представляют собой чрезвычайно реакционноспособные вещества. Их основным субстратом в реакциях свободнорадикального окисления являются ненасыщенные жирные кислоты, входящие в состав фосфолипидов мембран клетки (так называемые реакции перекисного окисления липидов). Интенсификация этих реакций может вызвать повреждение мембраны клетки, ее барьерной, рецепторной и обменной функций, модификацию молекул нуклеиновых кислот и белков, что ведет к мутациям и инактивации ферментов.
В физиологических условиях интенсификация перекисного окисления липидов регулируется антиоксидазной системой клеток, представленной ферментами, инактивируюшими активные формы кислорода — супероксиддисмутазой, каталазой, пероксидазой и веществами, обладающими антиокислительной активностью — токоферолом (витамин Е), убихиноном и др. Выраженный защитный эффект на мембраны клетки (цитопротекторный эффект) при различных повреждающих воздействиях на организм оказывают простагландины Е и J2, «гася» активацию свободнорадикального окисления. Простагландины защищают слизистую желудка и гепатоциты от химических повреждений, нейроны, клетки нейроглии, кардиомиоциты — от гипоксических повреждений, скелетные мышцы — при тяжелой физической нагрузке. Простагландины, связываясь со специфическими рецепторами на клеточных мембранах стабилизируют бислой последних, уменьшают потерю мембранами фосфолипидов.
Функции рецепторов мембран
text_fields
text_fields
arrow_upward
Химический или механический сигнал вначале воспринимается рецепторами мембраны клетки. Следствием этого является химическая модификация мембранных белков, влекущая активацию «вторичных посредников», обеспечивающих быстрое распространение сигнала в клетке к ее геному, энзимам, сократительным элементам и т.д.
Схематично трансмембранная передача сигнала в клетке может быть представлена следующим образом:
1) Возбужденный воспринятым сигналом рецептор активирует у — белки мембраны клетки. Это происходит при связывании ими гуанозинтрифосфата (ГТФ).
2) Взаимодействие комплекса «ГТФ-у- белки», в свою очередь, активирует фермент — предшественник вторичных посредников, расположенный на внутренней стороне мембраны.
• Предшественником одного вторичного посредника — цАМФ, образующегося из АТФ, является фермент аденилатциклаза;
• Предшественником других вторичных посредников — инозитолтрифосфата и диацилглицерина, образующихся из фосфатидилинозитол-4,5-дифосфата мембраны, является фермент фосфолипаза С. Кроме того, инозитолтрифосфат мобилизует в клетке еще один вторичный посредник — ионы кальция, участвующие практически во всех регуляторных процессах в клетке. Так, например, образовавшийся инозитолтрифосфат вызывает выброс кальция из эндоплазматического ретикулума и повышение его концентрации в цитоплазме, тем самым включая различные формы клеточного ответа. С помощью инозитолтрифосфата и диацилглицерина регулируется функция гладких мышц и В-клеток поджелудочной железы ацетилхолином, передней доли гипофиза тиреогропин-релизинг фактором, ответ лимфоцитов на антиген и т.д.
• В некоторых клетках роль вторичного посредника выполняет цГМФ, образующийся из ГТФ с помощью фермента гуанилатциклазы. Он служит, например, вторичным посредником для натрийуретического гормона в гладких мышцах стенок кровеносных сосудов. цАМФ служит вторичным посредником для многих гормонов — адреналина, эритропоэтина и др. (глава 3).
Читайте также:
doctor-v.ru
4. Строение и функции клеточных мембран.
3. Основным свойством живых клеток является раздражимость, т. е. их способность реагировать изменением обмена веществ в ответ на действие раздражителей. Возбудимость — свойство клеток отвечать на раздражение возбуждением. К возбудимым относят нервные, мышечные и некоторые секреторные клетки. Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения).
Cвойства возбудимых тканей.
4 свойства: возбудимость, проводимость, рефрактерность, лабильность. Для мышечной ткани характерна также сократимость.
Возбудимость – способность ткани отвечать на раздражение изменением ряда своих свойств. Показатель возбудимости – порог раздражения. Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани.
Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения возбуждения.
Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения. При этом в ходе ответной реакции ткань перестает воспринимать раздражитель.
Лабильность – способность ткани генерировать определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимого раздражения. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность).
Сократимость – способность мышцы отвечать сокращением на раздражение.
Раздражитель – фактор, способный вызвать ответную реакцию возбудимых тканей. В условиях физиологического эксперимента в качестве раздражителя чаще всего используют электрический ток. Хронаксия – наименьший промежуток времени, в течение которого ток силой в 2 реобазы (пороговая сила раздражителя для электрического тока) вызывает в ткани возбуждение.
Различают два вида биологических реакций: специфические и неспецифические. Специфические реакции характерны для какой-то строго определенной ткани
Все многообразие раздражителей можно выделить в отдельные группы. Классификация раздражителей зависит от того, что берется за основу:
По своей природе раздражители бывают: 1) химические; 2) физические; 3) механические; 4) термические; 5) биологические.
1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
Строение и функции ионных каналов. Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал (диаметр 0,5—0,7 нм). Расчеты показывают, что суммарная площадь каналов занимает незначительную часть поверхности клеточной мембраны.
Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. Канал неоднороден по своим функциональным характеристикам, особенно это касается белковых структур, находящихся у входа в канал и у его выхода (так называемые воротные механизмы).
Пассивный транспорт
-. Простая диффузия
-. Осмос
— Диффузия ионов
— Облегченная диффузия
Активный транспорт
— Первично-активный транспорт
-. Вторично-активный транспорт
5. Потенциа́л поко́я (ПП) — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ .У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки.
Согласно теории Ю. Бернштейна, при возбуждении клетки её мембрана повреждается, и ионы К+ вытекают из клетки по концентрационному градиенту до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя. Это утверждение, относящееся скорее к потенциалу действия, было опровергнуто Ходжкином и Хаксли в 1939 году.
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.
Итак, мембранный потенциал покоя — это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
6. Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.
Фазы потенциала действия
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
Отрицательный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Положительный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
studfiles.net
Биология для студентов — 05. Структура, химический состав и функции мембран в растительной клетке
Клеточная мембрана (также цитолемма, плазмалемма, или плазматическая мембрана) — эластическая молекулярная структура, состоящая из белков и липидов. Отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды.
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
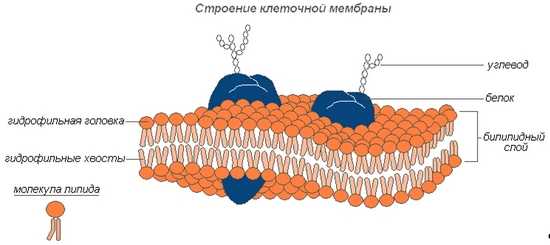
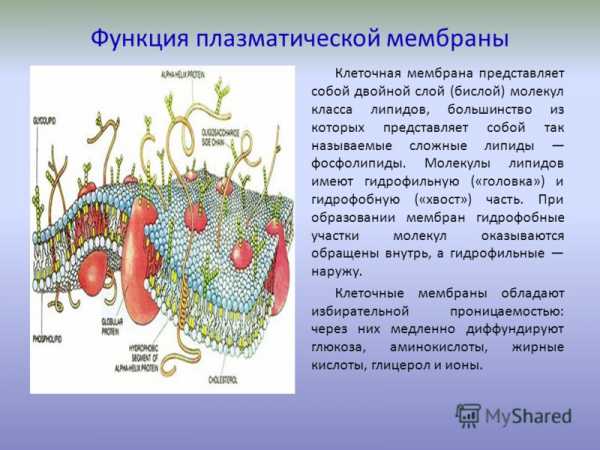
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») части. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Биологическая мембрана включает и различные белки:
- интегральные (пронизывающие мембрану насквозь),
- полуинтегральные (погружённые одним концом во внешний или внутренний липидный слой),
- поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны).
Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки и клеточной стенкой снаружи.
Функции мембран:
- Барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой.
- Транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
- Матричная — обеспечивает определённое взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- Механическая — обеспечивает автономность клетки, её внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечении механической функции имеют клеточные стенки.
- Энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки.
Мембраны состоят из липидов трёх классов:
- фосфолипиды,
- гликолипиды,
- холестерол.
Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой».
Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим— более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку.
Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются. Рядом с белками находятся аннулярные липиды — они более упорядочены, менее подвижны, имеют в составе более насыщенные жирные кислоты и выделяются из мембраны вместе с белком. Без аннулярных липидов белки мембраны не работают.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, в наружном содержатся преимущественно фосфатидилинозитол, фосфатидилхолин, сфингомиелины и гликолипиды, во внутреннем — фосфатидилсерин, фосфатидилэтаноламин и фосфатидилинозитол. Переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён, но может происходить спонтанно, примерно раз в 6 месяцев или с помощью белков-флиппаз и скрамблазыплазматической мембраны. Если в наружном слое появляется фосфатидилсерин, это является сигналом для макрофагов о необходимости уничтожения клетки.
Мембранные органеллы – это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
vseobiology.ru
Клеточные мембраны, их структура. Функции мембраны клетки.
Количество просмотров публикации Клеточные мембраны, их структура. Функции мембраны клетки. — 318
Таблица №2
Вопрос 1 (8)
Клеточная мембрана (или цитолемма, или плазмалемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды.
Функции клеточной или плазматической мембраны
Мембрана обеспечивает:
1) Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток; 2) Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала; 3) Специфику межклеточных контактов.
Благодаря наличию в мембране многочисленных рецепторов, воспринимающих химические сигналы — гормоны, медиаторы и другие биологически активные вещества, она способна изменять метаболическую активность клетки. Мембраны обеспечивают специфику иммунных проявлений, благодаря наличию на них антигенов — структур, вызывающих образование антител, способных специфически связываться с этими антигенами. Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных отсеках (компартментах) внутри клетки.
Структура мембраны клетки
Мембрана клетки — эластичная структура, толщиной от 7 до 11 нм (рис.1.1). Она состоит, в основном, из липидов и белков. От 40 до 90% всех липидов составляют фосфолипиды — фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, сфингомиелин и фосфатидилинозит. Важным компонентом мембраны являются гликолипиды, представленные церебризидами, сульфатидами, ганглиозидами и холестерином.
Основной структурой мембраны клетки является двойной слой фосфолипидных молекул. За счёт гидрофобных взаимодействий углеводные цепочки липидных молекул удерживаются друг возле друга в вытянутом состоянии. Группы же фосфолипидных молекул обоих слоев взаимно действуют с белковыми молекулами, погруженными в липидную мембрану. Благодаря тому, что большинство липидных компонентов бислоя находится в жидком состоянии, мембрана обладает подвижностью, совершает волнообразные движения. Ее участки, а также белки, погруженные в липидный бислой, перемешаются из одной ее части в другую. Подвижность (текучесть) мембран клеток облегчает процессы транспорта веществ через мембрану.
Белки мембраны клеток представлены, в основном, гликопротеинами.
Различают
интегральные белки, проникающие через всю толщу мембраны и
периферические белки, прикрепленные только к поверхности мембраны, в основном, к внутренней ее части.
Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и щелочная фосфатазы и др.). Но некоторые энзимы также представлены интегральными белками — АТФ-аза.
Интегральные белки обеспечивают селективный обмен ионов через каналы мембран между экстрацеллюлярной и интрацеллюлярной жидкостью, а также действуют как белки — переносчики крупных молекул.
Рецепторы и антигены мембраны бывают представлены как интегральными, так и периферическими белками.
Белки, примыкающие к мембране с цитоплазматической стороны, относятся к цитоскелету клетки. Οʜᴎ могут прикрепляться к мембранным белкам.
Так, белок полосы 3 (номер полосы при электрофорезе белков) эритроцитарных мембран объединяется в ансамбль с другими молекулами цитоскелета — спектрином через низкомолекулярный белок анкирин
Спектрин является основным белком цитоскелета͵ составляющим двумерную сеть, к которой прикрепляется актин.
Актин образует микрофиламенты, представляющие собой сократительный аппарат цитоскелета.
Цитоскелет позволяет клетке проявлять гибкоэластические свойства, обеспечивает дополнительную прочность мембраны.
Большинство интегральных белков — гликопротеины. Их углеводная часть выступает из клеточной мембраны наружу. Многие гликопротеины обладают большим отрицательным зарядом из-за значительного содержания сиаловой кислоты (к примеру, молекула гликофорина). Это обеспечивает поверхности большинства клеток отрицательный заряд, способствуя отталкиванию других отрицательно заряженных объектов. Углеводные выступы гликопротеинов являются носителями антигенов групп крови, других антигенных детерминант клетки, они действуют как рецепторы, связывающие гормоны. Гликопротеины образуют адгезивные молекулы, обуславливающие прикрепление клеток одна к другой, ᴛ.ᴇ. тесные межклеточные контакты.
Вопрос №2 (24)
referatwork.ru