Биологические мембраны
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ И ФУНКЦИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
Биологические мембраны — это активный молекулярный комплекс с высокоизбирательными свойствами, обеспечивающий обмен веществ и энергии с окружающей средой. В мембранах находятся специфические молекулярные насосы и каналы, с помощью которых регулируются молекулярный и ионный состав внутриклеточной среды. Помимо внешней цитоплазматической мембраны (плазмолемма) в клетках эукариотов имеются еще и внутренние мембраны, ограничивающие различные внутриклеточные компартменты (отсеки), например митохондрии, лизосомы, хлоропласты и т. д. Мембраны регулируют также обмен информацией между клетками и средой (восприятие внешних стимулов) и т. д. Мембраны различаются как по функции, так и по структуре. Однако всем им присущи следующие основные свойства:
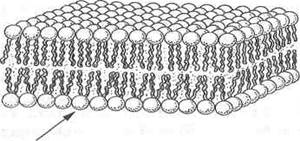
■ мембраны представляют собой плотную структуру толщиной в несколько молекул, 60-100 А, образующую сплошную перегородку между отдельными клетками и внутриклеточными отсеками;
■ мембраны главным образом состоят из липидов и белков. В мембранах имеются также углеводные компоненты, связанные с липидами и белками;
■ липиды мембран представлены относительно небольшими молекулами, несущими гидрофильные и гидрофобные группы. В водной среде эти молекулы спонтанно образуют замкнутые бимолекулярные слои, которые служат барьером для проникновения полярных соединений;
■ большинство функций мембран опосредуются специфическими белками, которые могут играть роль насосов, каналов, рецепторов, ферментов и т. д.
В состав мембран входят три основных типа липидов: фосфолипиды, гликолипиды и холестерин.
СТРОЕНИЕ МЕМБРАН
Фосфолипиды мембран. Среди липидных компонентов мембран главенствующая роль принадлежит фосфолипидам — веществам, производным либо трехатомного спирта глицерола (глицерофосфолипиды),
либо более сложного спирта сфингозина (сфингофосфолипиды). Все основные глицерофосфолипиды являются производными фосфатидной кислоты, этерифицированной с гидроксильной группой спиртов, таких как серии (серинфосфатиды — кефалины), этаноламин, холин (холинфосфа-тиды), кардиолипин (дифосфатидилглицерол) и инозитол (фос-фатидилинозитол).Из сфингофосфолипидов основным является сфингомиелин, основу которого составляет сфингозин — аминоспирт с длинной ненасыщенной углеводородной цепью. В состав сфингомиелина входит также азотистое основание холин.
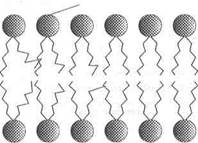
Независимо от структурных разнообразий каждая молекула фосфолипида в водной среде — это амфипатическая молекула с полярной головкой и неполярной хвостовой частью. Полярная головка образуется за счет остатков спиртовых групп, азотистых оснований и фосфорной кислоты. Хвостовая же часть — за счет радикалов двух жирных кислот насыщенного и ненасыщенного ряда. Благодаря своим амфипатическим свойствам фосфолипиды в водной среде спонтанно формируют липидные бислои, где полярные головки фосфолипидов направлены в сторону растворимой части клетки с образованием водородных связей с диполями воды, а неполярные хвосты — внутрь бислоя, скрепляясь между собой за счет гидрофобных взаимодействий. Именно бислойная структура фосфолипидов определяет полупроницаемые свойства мембран.
В качестве примера можно привести фосфатидилэтаноламин и фосфатидилхолин. Оба они имеют в верхней части молекулы полярные головки Nh5 (фосфатидилэтаноламин) и N+ (фосфатидилхолин), которые через остаток фосфорной кислоты и глицерина присоединены к двум остаткам жирных кислот, из которых одна насыщенная, другая — ненасыщенная (рис. 1).
|
|
|
|
|
|

|
Фосфолипиды с ненасыщенными жирными кислотами |
Фосфолипиды с насыщенными жирными кислотами |
В 1972 г. С. Дж. Сингер и Г. Никольсон сформулировали теорию строения мембран, согласно которой мембраны имеют жидкостно-мозаичную структуру. При обычной для клетки температуре мембранный бислой находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных фосфолипидов. Жирные кислоты с ненасыщенными связями характеризуются большей гибкостью (в отличие от насыщенных ЖК) и способностью создавать изгибы, что предотвращает плотную упаковку, затрудняет «замораживание» мембран и таким образом влияет на их текучесть ().
Упаковка углеводородов в бислое зависит от температуры. При низких температурах бислой находится в виде геля и упакован плотно, при высоких же температурах (температура тела) бислой фактически «расплавляется» и становится текучим, позволяя липидным молекулам двигаться вокруг своей оси, вращаться, меняться местами. Это, в свою очередь, способствует перемещению уже других компонентов в мембране, в частности белков.
Мембранные гликолипиды. Следующим важным компонентом мембран являются гликолипиды — липиды, содержащие углеводы. Гликолипиды животных клеток, подобно сфингомиелину, являются производными спирта сфингозина, связанного с ацильным радикалом. Отличие между этими липидами заключается в том, что в гликолипидах к сфингозиновому остатку присоединены один или несколько остатков сахара, а в сфингомиелине — фосфорилхолин.
Гликолипиды могут быть простые и сложные. Простейший гликолипид — цереброзид, содержащий только один остаток сахара (глюкозу или галактозу). В более сложных гликолипидах число сахарных остатков может достигать семи (ганглиозиды)
Гликолипиды в мембранах могут выполнять защитную, полупроводниковую, рецепторсвязывающую роль. Среди молекул, способных связываться с гликолипидами, встречаются также такие клеточные яды, как холера, токсин тетануса и др.
Холестерин в мембранах. Другой представитель липидов в мембранах — это холестерин. Количество его в мембранах варьирует в зависимости от типа клеток. В плазматических мембранах в среднем на каждую молекулу фосфолипида приходится примерно 1 молекула холестерина. У других (например, бактерий) — холестерина нет вообще. У холестерина так же, как у фосфолипидов, имеются участки полярные и неполярные.
Внутри мембран холестерин внедряется между фосфолипидами и ориентируется в том же направлении, что и сами молекулы фосфолипидов. Таким образом, полярная головка холестерина оказывается в той же плоскости, что и полярные головки фосфолипидов (рис. 2).
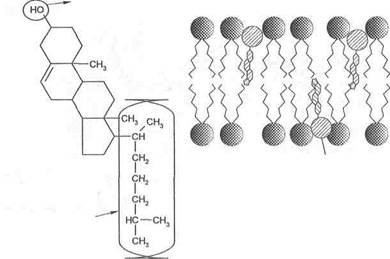
В мембранах холестерин выполняет следующие функции:
■ фиксируют первые несколько ближайших углеводородных групп, входящих в состав фосфолипидных жирных кислот. Это делает липидный бислой более устойчивым к деформациям и ограничивает прохождение через них небольших водорастворимых молекул. В случае отсутствия холестерина (как, например, у бактерий) клетка нуждается в оболочке;
■ предотвращает кристаллизацию углеводородов и фазовые сдвиги в мембране.
Мембранные белки. В то время как мембранные липиды ответственны за создание барьера проницаемости, мембранные белки опосредуют отдельные функции мембран, т. е. транспорт веществ, передачу информации, энергии и т. д. Соотношение между липидами и белками у разных мембран может быть разным, например, миелин, изолятор нервных клеток, содержит только 18% белков и 76% липидов, а митохондриальная внутренняя мембрана, наоборот — содержит 76% белков и только 24% липидов. В зависимости от характера локализации в мембранах выделяют белки интегральные (трансмембранные), периферические и «заякоренные».
Интегральные белки пронизывают бислой мембраны насквозь и благодаря своим бифильным свойствам фиксируются в нем. Белки, пронизывающие мембрану только один раз, называют однократно пронизывающими белками, а несколько раз — многократно пронизывающими.
Периферические
|
|
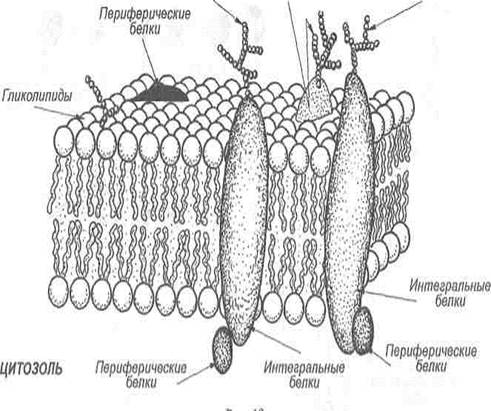
Олигосахариды Гликопротеины Олигосахариды
Рис. 3. Белковый состав мембран
«Заякоренные» белки фиксируются в мембранах с помощью коротких хвостовых липофильных доменов, образованных либо за счет гидрофобных аминокислотных остатков (цитохром b5), либо за счет ковалентно связанных ацильных радикалов (фермент щелочная фосфатаза).
Участки белков, которые обращены во внеклеточную среду, могут подвергаться гликозилированию.
Транспортные белки. Мембранным белкам принадлежит решающая роль в транспорте веществ через мембраны, и для выполнения этой роли наилучшим образом подходят интегральные белки, которые охватывают пространство как внутриклеточное, так и межклеточное.
Транспорт веществ через мембраны белки осуществляют различными способами; они могут выступать в качестве белковых насосов, каналов, транспортеров.
АТР — зависимые насосы, представляют собой АТРазы, которые способствуют движению через мембраны ионов или небольших молекул против их концентрационного градиента (или электрохимического потенциала) за счет энергии расщепления АТР. Такой вид транспорта известен как активный транспорт. С активным транспортом сопряжены определенные химические реакции, так, например, благодаря таким насосам в животных клетках обеспечивается поддержание низких концентраций Са2+ внутри клетки и высокое содержание ионов Nа+ в межклеточном пространстве, низкое значение рН в желудочном соке у человека и животных (моногастричных), внутри лизосом клеток, вакуолей растительных клеток.
Белковые каналы обеспечивают быстрое (до 108 молекул в секунду) перемещение одновременно молекул воды и других молекул и ионов по направлению снижения их концентрационного градиента (или электрохимического потенциала). Такие перемещения молекул обычно являются энергетически выгодными. Так, плазматические мембраны всех животных клеток содержат К+ — специфичные белковые каналы, которые открываются и закрываются в определенное время. Другие белковые каналы в это время закрыты и открываются только в ответ на воздействие специальных сигналов. Особенно большую роль играют такие каналы в нервных клетках.
Белки-транспортеры способствуют транспорту различных ионов и молекул через мембрану; однако, в отличие от канальных белков, белки-транспортеры связывают одну (или несколько) молекул субстрата одновременно, что приводит к изменению конформации белка и в результате к транспорту этих связанных молекул через мембрану. Такие транспортеры могут переносить в клетку около 102-104 молекул в секунду, что гораздо медленнее, чем движение по белковым каналам.
Обнаружены 3 типа белка-транспортера.
Юнипортеры осуществляют транспорт через мембрану животных клеток молекул одного типа в сторону уменьшения их концентрационного градиента, например, глюкозу, аминокислоты.
Антипортеры и симпортеры обеспечивают согласованный ко-транспорт одних молекул или ионов через мембрану против их концентрационного градиента с движением других молекул или ионов в процессе их перемещения в сторону уменьшения их концентрационного градиента.
АКТИВНЫЙ ТРАНСПОРТ ЧЕРЕЗ МЕМБРАНУ
Активный транспорт — это транспорт веществ через мембраны за счет потребления энергии расщепления АТР. Активным транспортом осуществляется транспорт некоторых ионов и небольших молекул против их концентрационного градиента.
Белки, участвующие в активном транспорте через мембраны (белковые насосы), условно подразделяют на 4 класса: суперсемейство белков АВС, белки класса Р., F., и V. Белки класса Р., F. и V транспортируют только ионы, а АВС — небольшие молекулы и ионы.
Белки (насосы) Р. — класса состоят из 2 субъединиц — α и β; α — субъединица содержит АТР — связывающий участок и является каталитической, а β — субъединица — регуляторной. Большинство белков этого класса являются тетрамерами, составленными из 2 α, и 2 β — субъединиц. В процессе транспорта, по крайней мере, одна из α — субъединиц сначала подвергается фосфорилированию (поэтому и обозначается как «Р»), и именно через нее происходит транспорт ионов.
К белкам Р — класса относятся:
■ Nа+/К+- АТРаза — фермент, локализованный в плазматической мембране и регулирующий внутриклеточное содержание ионов Nа+ и К+ в клетках животных;
■ Са2+- АТРазы — насосы, перекачивающие ионы Са2+ из цитозоля в межклеточное пространство против их концентрационного градиента для поддержания низкого уровня кальция (10-2 М) в цитоплазме клеток животных, дрожжей и растений. Помимо плазматических Са2+-АТРаз клетки мышц содержат еще другую Са2+-АТРазу (мышечный Са2+-й насос), которая осуществляет перекачивание ионов кальция из цитозоля в саркоплазматический ретикулум (СР) — внутриклеточное хранилище кальция;
■ мембранные белки эпителиальных клеток желудка у млекопитающих, способствующие поступлению соляной кислоты в желудок;
■ Н+- насосы, транспортирующие протоны водорода из клетки взамен поступления ионов К+ внутрь клетки;
■ Н+- насосы, регулирующие мембранный электрический потенциал в клетках растений, грибов, бактерий. Эти насосы не содержат фосфопротеиновой части.
Ионные насосы класса F и V структурно похожи друг на друга, но гораздо сложнее, чем белки класса Р. Насосы F и V состоят из 3 трансмембранных белков и 5 различных полипептидов, которые ориентированы в цитозольную часть белка и формируют внутрицитозольный домен. Некоторые субъединицы трансмембранных белков, ориентированные во внешнюю часть биомембран, структурно аналогичны внутрицитозольным доменным полипептидам.
Насосы класса V в основном участвуют в поддержании низкого значения рН в вакуолях растений и лизосомах и других кислотных везикулах животных клеток за счет расходования энергии расщепления АТР и перекачивая протоны водорода через мембрану из цитозоля в межклеточное пространство против протонного электрохимического градиента. Насосы класса F найдены в плазматических мембранах бактерий, мембранах хлоропластов и митохондрий. В отличие от насосов класса V их функция в основном направлена на синтез АТР из АDР и неорганического фосфата за счет движения протонов водорода из цитозольного межмембранного пространства в сторону уменьшения электрохимического градиента.
Последний класс АTР — зависимых транспортных белков — это суперсемейство АВС (АТР-binding cassette). Этот класс включает до 100 различных транспортных белков, и обнаружены они в клетках всех организмов. Каждый АВС — белок специфичен по отношению к одному какому-то субстрату, или группе субстратов, похожих друг на друга, включая ионы, углеводы, пептиды, полисахариды и даже белки.
Все АВС — транспортные белки объединяет наличие у них 4 главных доменов — двух трансмембранных доменов (Т), образующих так называемые ворота для «прохождения» молекул через мембрану, и двух внутрицитозольных домена (А), участвующих в связывании АТР. Таких АТР — связывающих участков у АВС — белков могут быть один или два, и их часто называют АТРазами, хотя и не всегда они проявляют АТР — гидролизующие свойства. В отдельных случаях такие трансмембранные белки могут проявить АТР — синтезирующие свойства, что играет решающую роль при синтезе АТР в митохондриальных мембранах.
Мембраны биологические — это… Что такое Мембраны биологические?
функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей.
Биологические мембраны имеются во всех клетках. Их значение определяется важностью функций, которые они выполняют в процессе нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при различных нарушениях мембранных функций и проявляющихся практически на всех уровнях организации — от клетки и субклеточных систем до тканей, органов и организма в целом. Мембранные структуры клетки представлены поверхностной (клеточной, или плазматической) и внутриклеточными (субклеточными) мембранами. Название внутриклеточных (субклеточных) мембран обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и др. (см. Клетка). Толщина биологических мембран — 7—10 нм, но их общая площадь очень велика, например, в печени крысы она составляет несколько сот квадратных метров. Химический состав и строение биологических мембран. Состав М.б. зависит от их типа и функций, однако основными составляющими являются Липиды и Белки, а также Углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса). Липиды. В составе М.б. обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды. В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит) и сфингофосфолипиды (производные церамида, сфингомиелин). Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%). В липидных компонентах М.б. содержатся разнообразные жирные кислоты, однако в мембранах животных клеток преобладают пальмитиновая, олеиновая и стеариновая кислоты. Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать двухслойные структуры (бислои) при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» —наоборот, наружу. Липидный бислои представляет собой динамичную структуру: образующие его липиды могут вращаться, двигаться в латеральном направлении и даже переходить из слоя в слой (флип-флоп переход). Такое строение липидного бислоя легло в основу современных представлений о структуре М.б. и определяет некоторые важные свойства М.б., например способность служить барьером и не пропускать молекулы веществ, растворенных в воде (рис.). Нарушение структуры бислоя может привести к нарушению барьерной функции мембран.Холестерин в составе М.б. играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов.
Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность.
Белки биологических мембран исключительно разнообразны. Молекулярная масса их в большинстве своем составляет 25 000 — 230 000.
Белки могут взаимодействовать с липидным бислоем за счет электростатических и (или) межмолекулярных сил. Они сравнительно легко могут быть удалены из мембраны. К такому типу белков относят цитохром с (молекулярная масса около 13 000), обнаруживаемый на наружной поверхности внутренней мембраны митохондрий.
Эти белки называются периферическими, или наружными. Для других белков, получивших название интегральных, или внутренних, характерно то, что одна или несколько полипептидных цепей оказываются погруженными в бислои или пересекают его, иногда не один раз (например, гликофорин, транспортные АТФ-азы, бактериородопсин). Часть белка, контактирующая с гидрофобной частью липидного бислоя, имеет спиральное строение и состоит из неполярных аминокислот, в силу чего между этими компонентами белков и липидов происходит гидрофобное взаимодействие. Полярные группы гидрофильных аминокислот непосредственно взаимодействуют с примембранными слоями, как с одной, так и с другой стороны бислоя. Молекулы белков, как и молекулы липидов, находятся в динамическом состоянии, для них также характерна вращательная, латеральная и вертикальная подвижность. Она является отражением не только их собственной структуры, но и функциональной активности. что в значительной степени определяется вязкостью липидного бислоя, которая, в свою очередь, зависит от состава липидов, относительного содержания и вида ненасыщенных жирно-кислотных цепей. Этим объясняется узкий температурный диапазон функциональной активности мембраносвязанных белков. Белки мембран выполняют три основные функции: каталитическую (ферменты), рецепторную и структурную. Однако такое разграничение достаточно условно, и в ряде случаев один и тот же белок может выполнять и репепторную и ферментную функции (например, инсулин). Число мембранных ферментов (Ферменты) в клетке достаточно велико, однако их распределение в различных типах М.б. неодинаково. Некоторые ферменты (маркерные) присутствуют только в мембранах определенного типа (например, Na, К-АТФ-аза, 5-нуклеотидаза, аденилатциклаза — в плазматической мембране; цитохром Р-450, НАДФН-дегидрогеназа, цитохром в5 — в мембранах эндоплазматического ретикулума; моноаминоксидаза — в наружной мембране митохондрий, а цитохром С-оксидаза, сукцинат-дегидрогеназа — во внутренней; кислая фосфатаза — в мембране лизосом). Рецепторные белки, специфически связывая низкомолекулярные вещества (многие гормоны, медиаторы), обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка принимает различные сигналы, поступающие из внешней среды.К структурным белкам относят белки цитоскелета, прилегающие к цитоплазматической стороне клеточной мембраны. В комплексе с микротрубочками и микрофиламентами цитоскелета они обеспечивают противодействие клетки изменению ее объема и создают эластичность. В эту же группу включают ряд мембранных белков, функции которых не установлены.
Углеводы в биологических мембранах находятся в соединении с белками (гликопротеины) и липидами (гликолипиды). Углеводные цепи белков представляют собой олиго- или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Углеводные компоненты М.б. открываются в основном во внеклеточную среду, образуя на поверхности клеточных мембран множество ветвистых образований, являющихся фрагментами гликолипидов или гликопротеидов. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обеспечением стабильности белковых молекул в М.б. Многие рецепторные белки содержат углеводные компоненты. Примером могут служить антигенные детерминанты групп крови, представленные гликолипидами и гликопротеинами. Функции биологических мембран. Барьерная функция. Для клеток и субклеточных частиц М.б. служат механическим барьером, отделяющим их от внешнего пространства. Функционирование клетки часто сопряжено с наличием значительных механических градиентов на ее поверхности преимущественно вследствие осмотического и гидростатического давления. Основную нагрузку в этом случае несет клеточная стенка, главными структурными элементами которой у высших растений являются целлюлоза, пектин и экстепин, а у бактерий — муреин (сложный полисахарид-пептид). В клетках животных необходимость в жесткой оболочке отсутствует. Некоторую жесткость этим клеткам придают особые белковые структуры цитоплазмы, примыкающие к внутренней поверхности плазматической мембраны. Перенос веществ через М.б. сопряжен с такими важнейшими биологическими явлениями, как внутриклеточный гомеостаз ионов, биоэлектрические потенциалы, возбуждение и проведение нервного импульса, запасание и трансформация энергии и т.п. (см. Биоэнергетика). Различают пассивный и активный транспорт (перенос) нейтральных молекул, воды и ионов через М.б. Пассивный транспорт не связан с затратами энергии, он осуществляется путем диффузии по концентрационным, электрическим или гидростатическим градиентам. Активный транспорт осуществляется против градиентов, связан с затратой энергии (преимущественно энергии гидролиза АТФ) и сопряжен с работой специализированных мембранных систем (мембранных насосов). Различают несколько видов транспорта. Если вещество транспортируется через мембрану независимо от наличия и переноса других соединений, то такой вид транспорта называют юнипортом. Если перенос одного вещества сопряжен с транспортом другого, то говорят о котранспорте, причем однонаправленный перенос называется симпортом, а противоположно направленный — антипортом. В особую группу выделяют перенос веществ путем экзо- и пиноцитоза. Пассивный перенос может осуществляться путем простой диффузии через липидный бислои мембраны, а также через специализированные образования — каналы. Путем диффузии через мембрану проникают в клетку незаряженные молекулы, хорошо растворимые в липидах, в т.ч. многие яды и лекарственные средства, а также кислород и углекислый газ. Каналы представляют собой липопротеиновые структуры, пронизывающие мембраны. Они служат для переноса определенных ионов и могут находиться в открытом или закрытом состоянии. Проводимость канала зависит от мембранного потенциала, что играет важную роль в механизме генерации и проведения нервного импульса. В ряде случаев перенос вещества совпадает с направлением градиента, но существенно превосходит по скорости простую диффузию. Этот процесс называют облегченной диффузией; он происходит с участием белков-переносчиков. Процесс облегченной диффузии не нуждается в энергии. Этим способом транспортируются сахара, аминокислоты, азотистые основания. Такой процесс происходит, например, при всасывании сахаров из просвета кишечника клетками эпителия. Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 106, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума — 104, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более 1/3 всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия — натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса — Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая — аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида. Изложенные механизмы транспорта различных веществ через клеточные мембраны имеют место и в случае их транспорта через эпителий ряда органов (кишечника, почек, легких), который осуществляется через слой клеток (монослой в кишечнике и нефронах), а не через единичную клеточную мембрану. Такой транспорт называют трансцеллюлярным, или трансэпителиальным. Характерной особенностью клеток, например эпителиоцитов кишечника и канальцев нефронов, является то, что апикальная и базальная их мембраны различаются по проницаемости, величине мембранного потенциала и транспортной функции. Способность генерировать биоэлектрические потенциалы и проводить возбуждение. Возникновение биоэлектрических потенциалов связано с особенностями строения биологических мембран и с деятельностью их транспортных систем, создающих неравномерное распределение ионов по обе стороны мембраны (см. Биоэлектрические потенциалы, Возбуждение). Процессы трансформации и запасания энергии протекают в специализированных М.б. и занимают центральное место в энергетическом обеспечении живых систем. Два основных процесса энергообразования — фотосинтез и тканевое дыхание — локализованы в мембранах внутриклеточных органелл высших организмов, а у бактерий — в клеточной (плазматической) мембране (см. Дыхание тканевое). Фотосинтезирующие мембраны преобразуют энергию света в энергию химических соединений, запасая ее в форме сахаров — основного химического источника энергии для гетеротрофных организмов. При дыхании энергия органических субстратов освобождается в процессе переноса электронов по цепи окислительно-восстановительных переносчиков и утилизируется в процессе фосфорилирования АДФ неорганическим фосфатом с образованием АТФ. Мембраны, осуществляющие фосфорилирование, сопряженное с дыханием, называют сопрягающими (внутренние мембраны митохондрий, клеточные мембраны некоторых аэробных бактерий, мембраны хроматофоров фотосинтезирующих бактерий). Метаболические функции мембран определяются двумя факторами: во-первых, связью большого числа ферментов и ферментативных систем с мембранами, во-вторых, способностью мембран физически разделять клетку на отдельные отсеки, отграничивая друг от друга метаболические процессы, протекающие в них. Метаболические системы не остаются при этом полностью изолированными. В мембранах, разделяющих клетку, имеются специальные системы, обеспечивающие избирательное поступление субстратов, выделение продуктов, а также движение соединений, обладающих регуляторным действием. Клеточная рецепция и межклеточные взаимодействия. Под этой формулировкой объединен весьма обширный и разнообразный набор важных функций клеточных мембран, определяющих взаимодействие клетки с окружающей средой и формирование многоклеточного организма как единого целого. Молекулярно-мембранные аспекты клеточной рецепции и межклеточных взаимодействий касаются прежде всего иммунных реакций, гормонального контроля роста и метаболизма, закономерностей эмбрионального развития. Нарушения структуры и функции биологических мембран. Разнообразие типов М.б., их полифункциональность и высокая чувствительность к внешним условиям порождают необыкновенное разнообразие структурно-функциональных нарушений мембран, возникающих при многих неблагоприятных воздействиях и сопряженных с огромным числом конкретных заболеваний организма как целого. Все это разнообразие нарушений достаточно условно можно подразделить на транспортные, функционально-метаболические и структурные. В общем виде охарактеризовать последовательность возникновения этих нарушений не представляется возможным, и в каждом конкретном случае требуется детальный анализ для выяснения первичного звена в цепи развития структурно-функциональных нарушений мембран. Нарушение транспортных функций мембран, в частности увеличение проницаемости мембран, — общеизвестный универсальный признак повреждения клетки. Нарушением транспортных функций (например, у человека) обусловлено более 20 так называемых транспортных болезней, среди которых почечная глюкозурия, цистинурия, нарушение всасывания глюкозы, галактозы и витамина В12, наследственный сфероцитоз и др. Среди функционально-метаболических нарушений М.б. центральными являются изменения процессов биосинтеза, а также многообразные отклонения в энергообеспечении живых систем. В наиболее общем виде следствием этих процессов является нарушение состава и физико-химических свойств мембран, выпадение отдельных звеньев метаболизма и его извращение, а также снижение уровня жизненно важных энергозависимых процессов (активного транспорта ионов, процессов сопряженного транспорта, функционирования сократительных систем и т.д.). Повреждения ультраструктурной организации М.б. выражаются в чрезмерном везикулообразовании, увеличении поверхности плазматических мембран за счет образования пузырей и отростков, слиянии разнородных клеточных мембран, образовании микропор и локальных структурных дефектов. Библиогр.: Биологические мембраны, под ред. Д.С. Парсонса, пер. с англ., М., 1978; Болдырев А.А. Введение в биохимию мембран, М., 1986, библиогр.; Конев С.В. и Мажуль В.М. Межклеточные контакты. Минск, 1977; Кульберг А.Я. Рецепторы клеточных мембран, М., 1987, библиогр.; Маленков А.Г. и Чуич Г.А. Межклеточные контакты и реакции ткани, М., 1979; Сим Э. Биохимия мембран, пер. с англ., М., 1985, библиогр.; Финеан Дж., Колмэн Р. и Митчелл Р. Мембраны и их функции в клетке, пер. с англ., М., 1977, библиогр.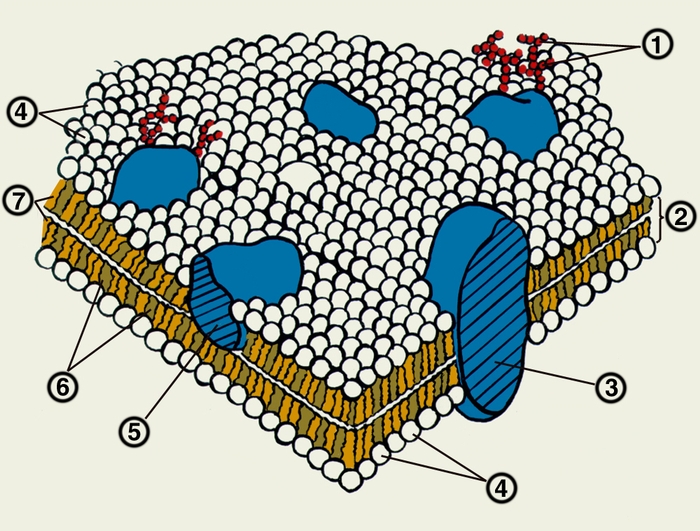 холестерин; 7 — жирнокислотные «хвосты» фосфолипидов»>
холестерин; 7 — жирнокислотные «хвосты» фосфолипидов»>Схема строения биологической мембраны: 1 — углеводные фрагменты гликопротеидов; 2 — липидный бислой; 3 — интегральный белок; 4 — «головки» фосфолипидов; 5 — периферический белок; 6 — холестерин; 7 — жирнокислотные «хвосты» фосфолипидов.
Биологическая мембрана — это… Что такое Биологическая мембрана?
- У этого термина существуют и другие значения, см. Мембрана
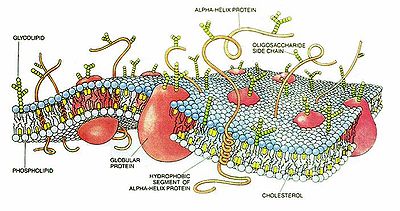 Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликс
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликсКле́точная мембра́на (или цитолемма, или плазмолемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая ее целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определенные условия внутриклеточной среды.
Основные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7-8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции биомембран
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетке воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
- маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс-одни вещества пропускают, а другие нет. существует четыре основных механизма для поступления веществ в клетку или их из клеки наружу:диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т.е. не требуют затрат энергии; два последних-активные процессы, связанные с потреблением энерги.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
Ссылки
- Bruce Alberts, et al. Molecular Biology Of The Cell. — 5th ed. — New York: Garland Science, 2007. — ISBN 0-8153-3218-1 — учебник по молекулярной биологии на англ. языке
- Рубин А.Б. Биофизика, учебник в 2 тт.. — 3-е издание, исправленное и дополненное. — Москва: издательство Московского университета, 2004. — ISBN 5-211-06109-8
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). — 1-е издание. — Москва: Мир, 1997. — ISBN 5-03-002419-0
- Иванов В.Г., Берестовский Т.Н. Липидный бислой биологических мембран. — Москва: Наука, 1982.
- Антонов В.Ф., Смирнова Е.Н., Шевченко Е.В. Липидные мембраны при фазовых переходах. — Москва: Наука, 1994.
См. также

Wikimedia Foundation. 2010.
Мембраны биологические — это… Что такое Мембраны биологические?
- У этого термина существуют и другие значения, см. Мембрана
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликсКле́точная мембра́на (или цитолемма, или плазмолемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая ее целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определенные условия внутриклеточной среды.
Основные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7-8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции биомембран
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетке воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
- маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс-одни вещества пропускают, а другие нет. существует четыре основных механизма для поступления веществ в клетку или их из клеки наружу:диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т.е. не требуют затрат энергии; два последних-активные процессы, связанные с потреблением энерги.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
Ссылки
- Bruce Alberts, et al. Molecular Biology Of The Cell. — 5th ed. — New York: Garland Science, 2007. — ISBN 0-8153-3218-1 — учебник по молекулярной биологии на англ. языке
- Рубин А.Б. Биофизика, учебник в 2 тт.. — 3-е издание, исправленное и дополненное. — Москва: издательство Московского университета, 2004. — ISBN 5-211-06109-8
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). — 1-е издание. — Москва: Мир, 1997. — ISBN 5-03-002419-0
- Иванов В.Г., Берестовский Т.Н. Липидный бислой биологических мембран. — Москва: Наука, 1982.
- Антонов В.Ф., Смирнова Е.Н., Шевченко Е.В. Липидные мембраны при фазовых переходах. — Москва: Наука, 1994.
См. также
Wikimedia Foundation. 2010.
Биологические мембраны — это… Что такое Биологические мембраны?
- У этого термина существуют и другие значения, см. Мембрана
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии — гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны — молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны — цепочки олигосахаридов, формирующие гликокаликсКле́точная мембра́на (или цитолемма, или плазмолемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая ее целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определенные условия внутриклеточной среды.
Основные сведения
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7-8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции биомембран
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетке воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
- маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс-одни вещества пропускают, а другие нет. существует четыре основных механизма для поступления веществ в клетку или их из клеки наружу:диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т.е. не требуют затрат энергии; два последних-активные процессы, связанные с потреблением энерги.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
Ссылки
- Bruce Alberts, et al. Molecular Biology Of The Cell. — 5th ed. — New York: Garland Science, 2007. — ISBN 0-8153-3218-1 — учебник по молекулярной биологии на англ. языке
- Рубин А.Б. Биофизика, учебник в 2 тт.. — 3-е издание, исправленное и дополненное. — Москва: издательство Московского университета, 2004. — ISBN 5-211-06109-8
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). — 1-е издание. — Москва: Мир, 1997. — ISBN 5-03-002419-0
- Иванов В.Г., Берестовский Т.Н. Липидный бислой биологических мембран. — Москва: Наука, 1982.
- Антонов В.Ф., Смирнова Е.Н., Шевченко Е.В. Липидные мембраны при фазовых переходах. — Москва: Наука, 1994.
См. также
Wikimedia Foundation. 2010.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ • Большая российская энциклопедия
БИОЛОГИ́ЧЕСКИЕ МЕМБРА́НЫ (лат. membrana – кожица, оболочка, перепонка), структуры, ограничивающие содержимое клеток (клеточная, или плазматическая, мембрана, плазмалемма) и внутриклеточных органелл. У прокариот имеется только клеточная мембрана, в большинстве случаев окружённая клеточной стенкой. У эукариот мембраной окружена не только клетка, но и ядро, а также митохондрии, лизосомы, пероксисомы, секреторные гранулы, эндосомы, у растений ещё – хлоропласты и вакуоли; мембраны образуют также разветвлённую сеть эндоплазматического ретикулума и комплекса Гольджи. Митохондрии, хлоропласты и ядра окружены двумя мембранами, а внутри хлоропластов имеется ещё один тип мембран, формирующих тилакоиды. У животных к клеточной мембране снаружи примыкает гликопротеиновый комплекс – гликокаликс, у растений – клеточная стенка. Толщина мембран варьирует от 6 до 10 нм.
Структура биологических мембран

Схема строения клеточной мембраны.
Основу Б. м. составляет протяжённый двойной слой (бислой) глицерофосфо-, сфинго- и гликолипидов со встроенными в него молекулами различных белков. Гидрофобные (неполярные) группы молекул липидов (остатки жирных кислот) погружены в толщу мембраны, а гидрофильные (полярные) головки ориентированы наружу, в окружающую водную среду (см. Липиды). Плотность упаковки Б. м. обеспечивается электростатическими взаимодействиями полярных головок и гидрофобными контактами между цепями жирных кислот. Входящие в состав Б. м. белки взаимодействуют с липидным бислоем с помощью гидрофобных взаимодействий и вандерваальсовых связей. Соотношение липидов и белков, их состав в разл. Б. м. могут существенно различаться. Так, в мембранах миелиновой оболочки содержание липидов (по массе) в четыре раза больше, чем белков, а во внутр. мембранах митохондрий более чем в два раза преобладают белки. Липиды Б. м. представлены гл. обр. фосфатидилхолином, фосфатидилэтаноламином, сфингомиелином, фосфатидилсерином, фосфатидилинозитом и кардиолипином, которые обнаруживаются примерно в одном и том же соотношении в мембранах разных по уровню организации организмов. В то же время набор жирных кислот, входящих в состав липидов, подвержен изменениям. Напр., понижение темп-ры, давления и солёности среды обитания организмов сопровождаются увеличением количества ненасыщенных связей и/или короткоцепочечных жирных кислот в фосфолипидах и происходящим вследствие этого уменьшением плотности упаковки бислоя. Липидам свойственна определённая подвижность внутри бислоя. Они способны к быстрому вращению вокруг оси (вращательная диффузия), к свободному перемещению в пределах одного слоя мембраны (латеральная диффузия), а также к переходу с одной стороны бислоя на другую (такое передвижение обеспечивается спец. механизмами). Для клеточных мембран животных клеток характерно высокое содержание холестерина (в ср. ок. 21%), который участвует в регуляции текучести мембраны, препятствуя плотной упаковке фосфолипидов. В растит. клетке роль холестерина играет его аналог – десмостерин. В мембранах бактерий и внутриклеточных органелл стерины отсутствуют. До 10% сухого вещества мембран приходится на долю углеводов, которые экспонированы на внешней стороне клеточной мембраны и являются составной частью мембранных гликолипидов и гликопротеинов.

Схема структурной организации интегрального белка. Спиральный участок встроен в гидрофобную часть липидного бислоя; N-концевой участок с прикреплёнными к нему олигосахаридными цепями расположен на вне…
Содержание белка в разл. мембранах колеблется от 20 до 75% (в пересчёте на сухую массу). Мембранные белки могут быть встроены в бислой (интегральные белки). При этом они погружены в мембрану и пронизывают её (иногда неск. раз) таким образом, что достаточно протяжённые участки белка, образованные гидрофобными аминокислотами, оказываются в её толще, а гидрофильные – на поверхности, по обе стороны Б. м. Выступающие над внешней стороной мембраны участки белковых молекул обычно несут неск. ковалентно связанных, часто разветвлённых цепей олигосахаридов, образованных остатками маннозы, фукозы, глюкозы, N-ацетилглюкозамина и др. Эти компоненты играют роль маркеров при распознавании клеточной поверхности. Молекулы периферических белков расположены гл. обр. на внутренней поверхности мембраны, не проникая внутрь бислоя, и удерживаются на ней с помощью электростатич. взаимодействий и водородных связей; они связываются с мембраной обратимо и могут переходить в цитоплазму при модификации белков (напр., путём их фосфорилирования) в ответ на изменения функционального состояния клетки. Мн. белки организованы в виде сложных комплексов (напр., белки дыхательной цепи митохондрий). В клетках происходит постоянное обновление компонентов Б. м. путём введения новых молекул липидов и белков, однако структурная организация Б. м. в течение всей жизни клетки остаётся неизменной.
Функции мембран
Осн. функции мембран связаны с белками. Мн. мембранные белки – ферменты, обеспечивающие протекание окислительно-восстановительных, гидролитич. и биосинтетич. реакций как на поверхности мембраны, так и внутри неё. Важнейшая функция белков мембран – транспортная. Жирорастворимые соединения (напр., стероидные гормоны) легко проникают в липидный бислой, но для большинства др. соединений (в т. ч. аминокислот, сахаров) и неорганич. ионов он непроницаем. Специально предназначенные для этих целей мембранные белки обеспечивают как активный (требующий затрат энергии), так и пассивный (за счёт градиента концентраций) транспорт веществ и ионов (см. Ионные каналы). Специфич. белки – ионные насосы ($\ce{Na/K}$-насос и $\ce{Ca}$-насос) ответственны за асимметричное распределение ионов $\ce{Na^+, K^+}$ и $\ce{Ca^{2+}}$ по обе стороны клеточной мембраны (напр., в цитоплазме животной клетки – низкую для $\ce{Na^+}$ и $\ce{Ca^{2+}}$ и высокую для $\ce{K^+}$). Такая асимметрия обеспечивает мн. проявления жизнедеятельности (электровозбудимость, осмотическую устойчивость и др.). Мембранные белки аквапорины образуют в мембране спец. каналы, регулирующие проникновение в клетку молекул воды. Взаимодействие клетки с внешней средой, регуляция внутриклеточных процессов осуществляются посредством рецепторных белков (рецепторов), ответственных за фото-, термо-, механо- и хеморецепцию.
Барьерная функция Б. м. обеспечивает сохранение определённого состава клетки и концентрации составляющих её веществ, а также защиту от воздействия разл. чужеродных факторов и токсинов. Благодаря Б. м. внутри клеток возможно одноврем. протекание множества несовместимых друг с другом реакций. Напр., необходимые клетке белки синтезируются на рибосомах, прикреплённых к эндоплазматическому ретикулуму, а их распад происходит в лизосомах. В Б. м. протекают процессы энергообмена клеток. Внутр. мембраны митохондрий и мембраны тилакоидов – важнейшие преобразователи энергии, играющие ключевую роль в запасании энергии, образующейся в ходе дыхания и фотосинтеза, в энергию пирофосфатной связи аденозинтрифосфата. Б. м. нейронов могут генерировать и осуществлять передачу электрич. сигнала, участвуя тем самым в процессах возбуждения и проведения нервного импульса.
Белковые и липидные компоненты выполняют ряд др. функций. Фрагменты фосфолипидов могут выступать в качестве предшественников сигнальных молекул (мессенджеров). Напр., при активации мембранной фосфолипазы А из бислоя высвобождается арахидоновая кислота, дальнейшие превращения которой приводят к образованию биологич. регуляторов – тромбоксанов, лейкотриенов и простагландинов. Фосфатидилсерин, локализованный на внутр. стороне мембраны, при инициации апоптоза мигрирует на её внешнюю сторону. Его появление служит сигналом для фагоцитов, которые имеют рецепторы на этот фосфолипид; они «узнают» дефектные клетки и уничтожают их. Гликолипиды наряду с гликопротеинами играют важную роль в явлениях межклеточной адгезии, участвуют в иммунных реакциях.
Методы изучения биологических мембран
Для изучения структуры и функции мембран используются электрофизиологич. и иммуноцитохимич. методы, жидкостная хроматография (для идентификации и анализа липидных компонентов), проточная цитометрия, позволяющая проследить ответ клетки на взаимодействие специфич. лигандов с клеточной мембраной, разнообразные физич. методы, характеризующие структуру мембран, упаковку и подвижность липидов в бислое (в т. ч. электронная микроскопия, малоугловое рассеивание нейтронов, флуоресцентная спектроскопия, круговой дихроизм), и др. методы.
Разнообразие типов Б. м., их полифункциональность и высокая чувствительность к внешним воздействиям являются причиной того, что они вовлекаются в разл. патологич. процессы. Повреждения клеточных мембран, приводящие к образованию свободных радикалов и гибели нервных клеток, лежат в основе нейродегенеративных заболеваний (болезнь Альцгеймера, паркинсонизм, боковой амиотрофический склероз), могут служить причиной инсульта и инфаркта миокарда.
ОСНОВНЫЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ БИОМЕМБРАН. «БИОЛОГИЧЕСКАЯ ХИМИЯ», Березов Т.Т., Коровкин Б.Ф.
ОСНОВНЫЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ БИОМЕМБРАН
Все клетки отграничены друг от друга и от окружающей среды с помощью специальной оболочки – клеточной мембраны. Со времен К. Негели, описавшего в 1855 г. структуру мембран, окружающих живые клетки, представления об устройстве и функциях мембран существенно обогатились. Клеточная мембрана во многом определяет свойства, поведение и даже форму клетки. Мембраны прокариот и эукариот различаются между собой по составу и свойствам. Растительные и животные клетки также отличаются друг от друга как по набору органелл, так и по свойствам мембран (рис. 9.1).
Состав и строение биологических мембран. Биологические мембраны состоят из белков и липидов. Углеводы присутствуют лишь в качестве составных частей сложных белков (гликопротеинов) и сложных липидов (гликолипидов). Нуклеиновые кислоты в небольшом количестве бывают ассоциированы с мембранами, но в состав мембранных структур не включаются. Вода составляет 20% от мембранного материала, а отношение белок/липид в зависимости от вида мембран колеблется от 0,25 (клетки миелиновой оболочки) до 3,0 (митохондриальные мембраны).
Липиды мембран представлены четырьмя основными группами: фосфо-липидами (основная доля), сфинголипидами, гликолипидами и стероидами. Фосфолипиды – это сложные эфиры фосфатидной кислоты. Основными фосфолипидами являются фосфатидилэтаноламин, фосфатидилсерин, фос-фатидилинозит и фосфатидилхолин. В состав фосфолипидов входят также дифосфатидилглицерины (кардиолипин), плазмалогены (1-О-алкенил-2-О ацилфосфолипиды) и диольные фосфолипиды. Сфинголипиды, которые являются производными церамида и монофосфорных эфиров различных спиртов, представлены в основном сфингомиелином. Гликолипиды – глико-зильные производные церамида – представлены как нейтральными церебро-зидами, так и их кислыми сульфоэфирами – сульфатидами. Производные церамида и нейраминовой кислоты – ганглиозиды – часто выделяют в отдельную группу липидов – гликосфинголипиды. Стероиды представлены холестерином (в мембранах животных клеток), ситостерином (в растительных клетках) и тетрахименином (обнаружен у тетрахимены).
Несмотря на различия в составе, все мембранные липиды построены по единому плану и легко смешиваются друг с другом, образуя монослойные или бислойные структуры (рис. 9.2). В этих структурах реализуется 2 типа взаимодействий: ионные взаимодействия полярных «голов» и гидрофобные взаимодействия жирнокислотных цепей. Благодаря этому мицеллы и липо-сомы, создаваемые протяженными бислойными структурами, достаточно стабильны в водном окружении.
В наружных (плазматических) мембранах животных клеток обнаруживается большое количество холестерина (около 21 моль%), меньше – фосфатидилэтаноламина и еще меньше фосфатидилхолина. Для внутриклеточных мембран основным компонентом является фосфатидилхолин, и соотношение фосфатидилхолин/фосфатидилэтаноламин в них всегда больше 1.
Рис. 9.1. Схематическое изображение животной (слева) и растительной (справа) клетки.
Соотношение основных классов липидов мембран нейронов у различных животных почти не подвержено изменениям. По-видимому, это соотношение сформировалось на самых ранних стадиях эволюции и обеспечивает как стабильность липидного бислоя, так и возможность включения в него белковых молекул. В то же время жирнокислотные компоненты мембранных липидов сильно подвержены эволюционной и сезонной изменчивости.
Рис. 9.2. Химические формулы распространенных липидов биологических мембран. 1 - фосфатидилэтаноламин; 2 — фосфатидилсерин; 3 — фосфатидилинозит; 4 - фосфатидилхолин; 5 — кардиолипин; 6 — сфингомиелин; 7 — цереброзид; 8 - холестерин; 9 — расположение молекулы холестерина между двумя молекулами фосфолипидов: а — наименее упорядоченная область бислоя, б — область, упорядочиваемая холестерином, в — область полярных «голов». Красным отмечены полярные области молекул.
Жирные кислоты, составляющие «хвост» липидных молекул, представлены насыщенными [от лауриновой (С12) до лигноцериновой (С24)] и ненасыщенными (мононенасыщенные пальмитоолеиновая и олеиновая; полиненасыщенные линолевая, линоленовая, арахидоновая) кислотами. У высших растений преобладают пальмитиновая, олеиновая и линолевая кислоты, а стеариновая почти не обнаруживается; в ряде случаев выявляются оксикислоты. В мембранах животных клеток, кроме пальмитиновой и олеиновой, много стеариновой кислоты и больше высокомолекулярных жирных кислот (содержат 20–24 углеродных фрагмента). Жирные кислоты, как правило, имеют четное число атомов углерода, но у цереброзидов и ган-глиозидов встречаются и нечетные углеводородные остовы. У бактерий полиненасыщенные жирные кислоты практически отсутствуют, но часто имеются разветвленные окси- и циклопропансодержащие кислоты. Для мембран термоацидофильных, галофильных и метанообразующих архебак-терий характерно наличие нетипичных липидов, содержащих изопреноидные цепи, метальные концы которых соединены друг с другом ковалент-ными связями. Такие «шпильки» обеспечивают повышенную прочность липидного бислоя. (Подробнее о жирных кислотах см. главу 11.)
Рис. 9.3. Распределение липидов между наружной (а) и
внутренней (б) сторонами бислоя в мембранах эритроцитов (I), вируса гриппа (II) и саркоплазматического
ретикулума кролика (III). Фл — общие
фосфолипиды; Фх — фосфатидилхолин; Фэ — фосфатидилэтаноламин; Фс -
фосфати-дилсерин; См — сфингомиелин; Фи — фосфатидилинозит.
Липиды в составе бислоя распределяются асимметрично. Это свойство диктуется особенностями строения их молекул: фосфатидилхолину, фосфат-идилсерину, сфингомиелину присуща цилиндрическая форма, фосфатидил-этаноламину – форма конуса, а лизофосфолипидам (получаются в результате отщепления от молекулы одной жирнокислотной цепи) – форма перевернутого конуса. Природные мембраны также обладают исходной асимметрией (рис. 9.3).
Белки взаимодействуют с мембранным бислоем, в результате чего они либо ассоциируются с поверхностью мембраны – периферические белки, либо пересекают бислой один или несколько раз, прочно интегрируясь в него,– это интегральные белки. Интеграция оказывается возможной, если в первичной структуре белка имеются достаточно протяженные участки, содержащие гидрофобные аминокислотные последовательности. В таком случае белковые молекулы способны самопроизвольно встраиваться в би-слой. При ассоциации рибосом с мембранными структурами встраивание гидрофобных белков в мембрану осуществляется синхронно с их синтезом при участии специальных механизмов, потребляющих энергию АТФ.
Участки белка, которые обращены во внеклеточную среду, могут подвергаться гликозилированию. В мембранах растений и бактерий полисахара играют самостоятельную роль, образуя наружную оболочку. В клетках животных, в которых наружный слой включает углеводы, имеется внутренний цитоскелет, состоящий из актина и других легко полимеризующихся белков; он имеет регулярную связь с мембранными белками и выполняет формообразующую и опорную функцию (рис. 9.4).
Фазовое состояние мембранных липидов. Мембранные липиды могут находиться в нескольких фазовых состояниях, т. е. они обладают мезомор-физмом. Два основных ламеллярных состояния, характерных для мембранных липидов в клеточных системах: кристаллическое и жидкокристаллическое – различаются плотностью упаковки и подвижностью находящихся в бислое белковых молекул. При более плотной упаковке ацильные цепи липидов расположены под углом, близким к 90°, и все С—С-связи находятся в транс-конформации (максимально вытянуты). Фазовый переход приводит к увеличению подвижности ацильных цепей в бислое, увеличению угла их наклона и уменьшению плотности упаковки. Латеральная подвижность мембранных белков после фазового перехода возрастает, увеличивается вероятность образования их ассоциатов.
Рис. 9.4. Схематическое изображение клеточной мембраны.
В липидном бислое могут также образовываться гексагональные структуры (вывернутые мицеллы). При их образовании в мембране возникают дефекты регулярной упаковки, что позволяет проникать через мембрану крупным молекулам, а также обеспечивает обмен компонентами монослоев в бислойной мембране.
Фазовые переходы мембранных липидов могут быть вызваны изменением температуры среды. Значение температуры, при котором наблюдается фазовый переход, называется критической температурой фазового перехода, или разделения фаз, если различные участки мембраны вследствие гетерогенности липидного состава по-разному отвечают на изменения температуры. Ионы Са2+, изменение числа ненасыщенных жирнокислотных цепей мембранных фосфолипидов и некоторые другие факторы также могут индуцировать фазовые переходы в бислое. Обычно критическая температура фазовых переходов приближена к температуре тела гомойотермных животных (или к температуре среды обитания пойкилотермных животных). Таким образом, достаточно незначительного изменения условий, чтобы изменить упаковку мембраны.
Специфические свойства биологических мембран. Благодаря указанным особенностям биологические мембраны имеют присущие им характерные черты. Они образуют протяженные бислойные структуры малой толщины (6–10 нм), объединяющие белковые и липидные компоненты с различными свойствами.
Целостная структура мембраны создается за счет гидрофобных и электростатических взаимодействий, а не за счет ковалентных связей между составляющими ее молекулами белков и липидов. Гидрофобный липидный бислой представляет естественную преграду для проникновения полярных молекул. Мембраны асимметричны по своему исходному строению, что обеспечивает градиент кривизны и спонтанное образование замкнутых структур.
Мембранный бислой обладает относительно малой микровязкостью. Другими словами, мембраны рыхло упакованы, что позволяет отдельным компонентам проявлять высокую подвижность в латеральном направлении.
Наружные мембраны клеток отличаются от внутренних по липидному составу (последние почти не содержат стеринов, имеют соотношение ФХ/ФЭ > 1) и обладают специфическим набором ферментов и рецепторов. Как правило, белки плазматических мембран со стороны внеклеточной среды обильно гликозилированы. Внутриклеточные мембраны содержат мало гликопротеинов и гликолипидов и характеризуются меньшей микровязкостью. Благодаря этому они могут образовывать органеллы малого размера. Мембранные белки выполняют различные специфические функции: рецепторные, транспортные, ферментативные, энергопреобразующие и т.д. (см. далее).
Функции биологических мембран. Как отмечалось, клеточные мембраны отграничивают содержимое клетки (или клеточной органеллы) от окружающей среды. Благодаря наличию специальных рецепторов они воспринимают сигналы из внешней среды (например, молекулы гормонов, называемые первичными мессенджерами, или посредниками), в ответ на которые образуются вторичные мессенджеры, высвобождающиеся внутрь клетки. Так осуществляется преобразование сигналов, изменяющих клеточный метаболизм в соответствии с изменяющимися условиями среды (см. главу 8).
Мембранные рецепторы выполняют функции узнавания (иммунокомпе-тентная система), адгезии (обеспечение межклеточных контактов, формирование тканей), регуляции активности ионных каналов (электрическая возбудимость, создание мембранного потенциала). Мембранные ферменты в составе бислоя приобретают большую стабильность и способность к осуществлению реакций, которые в гидрофильном окружении протекали бы с весьма малой скоростью. Липидное окружение предоставляет таким белкам «привилегированные» условия функционирования, но и накладывает ограничения на поведение белковых ассоциатов: последнее сильно зависит от плотности упаковки (микровязкости) мембран. Поэтому факторы, влияющие на липидный состав и свойства клеточной мембраны, оказывают регулирующее влияние на функции мембранных белков.
Мембранные белки часто образуют олигомерные ансамбли, взаимодействия между которыми (или длительность их существования в бислое) оказывается под контролем их мембранного окружения. Изменения микровязкости мембран в таком случае позволяют контролировать активность этих надмолекулярных структур.
Транспортная функция является одной из важных функций клеточных мембран (рис. 9.5). Мембрана создает существенные ограничения для проникновения различных веществ, однако она не является полностью непроницаемой: небольшие нейтральные молекулы могут проникать через бислой в области структурных дефектов. Этот процесс осуществляется по градиенту концентрации переносимого вещества — из области, где его содержание высоко, в область с более низким содержанием. Такой процесс называется простой диффузией, он осуществляется неизбирательно и с низкой скоростью.
При облегченной диффузии вещества также переносятся в направлении их концентрационного градиента, но с использованием специальных структур — переносчиков или каналов, увеличивающих скорость и специфичность переноса. Известны высокоспецифические транслоказы - белковые молекулы, переносящие аденило-вые нуклеотиды через внутреннюю мембрану митохондрий: Nа+/Са2+-обменник -белок, входящий в состав плазматических мембран многих клеток; низкомолекулярный пептид бактериального происхождения валиномицин — специфический переносчик для ионов К+.
Рис. 9.5. Перенос веществ через мембрану. а — виды переноса; б — пассивный и активный транспорт: 1 — пассивная диффузия; 2 - диффузия с помощью канала; 3 — диффузия с помощью переносчика; 4 — активный транспорт; 5 — вторично-активный транспорт.
Облегченная диффузия, осуществляемая с помощью каналов, не обладает высокой специфичностью (специфичность определяется лишь размерами канала), но протекает с большей скоростью, а процесс переноса не достигает насыщения в широком диапазоне концентраций переносимого вещества. Функционирование каналов в меньшей степени зависит от фазового состояния мембраны, чем функционирование переносчиков. Все эти примеры относятся к пассивному транспорту через мембрану.
Активный транспорт веществ осуществляется такими же механизмами, но протекает против концентрационного градиента и для своего осуществления должен быть сопряжен с энергодающим процессом. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы. Примером систем активного транспорта ионов является Na+/K+-АТФаза плазматических мембран животных клеток, которая «выкачивает» из клетки ионы натрия в обмен на ионы калия, затрачивая на выполнение этой работы АТФ в стехиометрии 3Nа+/2К+/1АТФ. Са2+-АТФаза осуществляет активный транспорт кальция через мембрану со стехиометрией 2Са2+/1АТФ.
В так называемых сопрягающих мембранах имеются протонные насосы, работающие как Н+-АТФазы. В результате их функционирования на мембране
возникает разность концентраций протонов (ΔрН) и разность электрических потенциалов, в совокупности образующие протонный электрохимический потенциал, обозначаемый ΔμН+ (см. далее). За счет работы Н+-АТФазы создается кислая среда в некоторых органеллах клетки (например, лизосомах, хромаффинных клетках надпочечников). В митохондриальной мембране Н+-АТФаза работает в обратном направлении, используя ΔμН+, создаваемый в дыхательной цепи, для образования АТФ.
Наконец, в клетках широко представлен вторично-активный транспорт, в процессе которого градиент одного вещества используется для транспорта другого. С помощью вторично-активного транспорта клетки аккумулируют сахара, аминокислоты и выводят некоторые продукты метаболизма, используя градиент Na+, создаваемый в ходе работы Na+/К+-АТФазы (см. рис. 9.5).
Предыдущая страница | Следующая страница
СОДЕРЖАНИЕ
мембрана | Определение, структура и функции
Мембрана , в биологии, тонкий слой, который образует внешнюю границу живой клетки или внутреннего компартмента клетки. Внешняя граница — это плазматическая мембрана, а отделы, окруженные внутренними мембранами, называются органеллами. Биологические мембраны выполняют три основные функции: (1) они не допускают попадания токсичных веществ в клетку; (2) они содержат рецепторы и каналы, которые позволяют определенным молекулам, таким как ионы, питательные вещества, отходы и продукты метаболизма, которые опосредуют клеточную и внеклеточную активность, проходить между органеллами и между клеткой и внешней средой; и (3) они разделяют жизненно важные, но несовместимые метаболические процессы, происходящие внутри органелл.
Молекулярный вид клеточной мембраны Внутренние белки проникают в липидный бислой и прочно связываются с ним, который состоит в основном из фосфолипидов и холестерина и который обычно составляет от 4 до 10 нанометров (нм; 1 нм = 10 −9 метров) ) по толщине. Внешние белки слабо связаны с гидрофильными (полярными) поверхностями, которые обращены к водной среде как внутри, так и снаружи клетки. Некоторые внутренние белки представляют собой боковые цепи сахара на внешней поверхности клетки. Encyclopdia Britannica, Inc.Британская викторина
Человеческое тело
Какова средняя температура здорового человека в градусах Цельсия?
Мембраны состоят в основном из липидного бислоя, который представляет собой двойной слой молекул фосфолипидов, холестерина и гликолипидов, содержащий цепочки жирных кислот и определяющий, образуется ли мембрана в виде длинных плоских листов или круглых пузырьков.Липиды придают клеточным мембранам жидкий характер с консистенцией, приближающейся к легкому маслу. Цепи жирных кислот позволяют многим маленьким жирорастворимым молекулам, таким как кислород, проникать через мембрану, но они отталкивают большие водорастворимые молекулы, такие как сахар, и электрически заряженные ионы, такие как кальций.
В липидный бислой встроены большие белки, многие из которых переносят ионы и водорастворимые молекулы через мембрану. Некоторые белки в плазматической мембране образуют открытые поры, называемые мембранными каналами, которые обеспечивают свободную диффузию ионов в клетку и из нее.Другие связываются с определенными молекулами на одной стороне мембраны и переносят молекулы на другую сторону. Иногда один белок одновременно транспортирует два типа молекул в противоположных направлениях. Большинство плазматических мембран содержат около 50 процентов белка по весу, в то время как мембраны некоторых метаболически активных органелл состоят на 75 процентов из белка. К белкам на внешней стороне плазматической мембраны прикреплены длинные молекулы углеводов.
различных типов мембранного транспорта Клеточная мембрана содержит белки, которые переносят ионы и водорастворимые молекулы внутрь или из клетки.Некоторые молекулы могут свободно диффундировать через мембрану в процессе, известном как простая диффузия. Encyclopædia Britannica, Inc.Многие клеточные функции, включая поглощение и преобразование питательных веществ, синтез новых молекул, производство энергии и регуляцию метаболических последовательностей, выполняются в мембранных органеллах. Ядро, содержащее генетический материал клетки, окружено двойной мембраной с большими порами, которые позволяют обмениваться материалами между ядром и цитоплазмой.Наружная ядерная мембрана является продолжением мембраны эндоплазматического ретикулума, которая синтезирует липиды для всех клеточных мембран. Белки синтезируются рибосомами, которые либо прикреплены к эндоплазматическому ретикулуму, либо свободно взвешены в содержимом клетки. Митохондрии, окисляющие и запасающие энергию единицы клетки, имеют внешнюю мембрану, легко проницаемую для многих веществ, и менее проницаемую внутреннюю мембрану, усеянную транспортными белками и производящими энергию ферментами.
Получите эксклюзивный доступ к контенту из нашего 1768 First Edition с подпиской. Подпишитесь сегодня ,клеточная мембрана | Определение, функции и структура
Клеточная мембрана , также называемая плазматической мембраной , тонкая мембрана, которая окружает каждую живую клетку, отделяя клетку от окружающей среды. Этой клеточной мембраной (также известной как плазматическая мембрана) окружены составляющие клетки, часто большие, водорастворимые, высоко заряженные молекулы, такие как белки, нуклеиновые кислоты, углеводы и вещества, участвующие в клеточном метаболизме. Вне клетки, в окружающей водной среде, находятся ионы, кислоты и щелочи, токсичные для клетки, а также питательные вещества, которые клетка должна поглощать, чтобы жить и расти.Таким образом, клеточная мембрана выполняет две функции: во-первых, быть барьером, удерживающим составляющие клетки внутри и нежелательными веществами, и, во-вторых, быть воротами, позволяющими переносить в клетку необходимые питательные вещества и перемещаться из клетки отходов. товары.
Молекулярный вид клеточной мембраны Внутренние белки проникают в липидный бислой и прочно связываются с ним, который состоит в основном из фосфолипидов и холестерина и обычно составляет от 4 до 10 нанометров (нм; 1 нм = 10 -9 метров) ) по толщине.Внешние белки слабо связаны с гидрофильными (полярными) поверхностями, которые обращены к водной среде как внутри, так и снаружи клетки. Некоторые внутренние белки представляют собой боковые цепи сахара на внешней поверхности клетки. Encyclopdia Britannica, Inc.Британская викторина
Человеческое тело
Где находится глотка?
Клеточные мембраны состоят в основном из липидов и белков на основе жирных кислот.Мембранные липиды в основном бывают двух типов: фосфолипиды и стерины (обычно холестерин). Оба типа разделяют определяющую характеристику липидов — они легко растворяются в органических растворителях, но, кроме того, у них обоих есть область, которая притягивается к воде и растворяется в ней. Это «амфифильное» свойство (обладающее двойным притяжением; то есть наличие как жирорастворимой, так и водорастворимой области) является основным для роли липидов как строительных блоков клеточных мембран. Мембранные белки также бывают двух основных типов.Один тип, называемый внешними белками, слабо прикреплен ионными связями или кальциевыми мостиками к электрически заряженной фосфорильной поверхности бислоя. Они также могут присоединяться ко второму типу белков, называемым внутренними белками. Внутренние белки, как следует из их названия, прочно встроены в фосфолипидный бислой. Как правило, мембраны, активно участвующие в метаболизме, содержат более высокую долю белка.
Химическая структура клеточной мембраны делает ее удивительно гибкой, идеальной границей для быстро растущих и делящихся клеток.Тем не менее, мембрана также является грозным барьером, позволяющим проходить одним растворенным веществам или растворенным веществам, блокируя другие. Липидорастворимые молекулы и некоторые небольшие молекулы могут проникать через мембрану, но липидный бислой эффективно отталкивает множество крупных водорастворимых молекул и электрически заряженных ионов, которые клетка должна импортировать или экспортировать, чтобы жить. Транспорт этих жизненно важных веществ осуществляется определенными классами внутренних белков, которые образуют множество транспортных систем: некоторые из них представляют собой открытые каналы, которые позволяют ионам диффундировать непосредственно в клетку; другие — «посредники», которые помогают растворенным веществам проходить сквозь липидный экран; а другие представляют собой «насосы», которые заставляют растворенные вещества проходить через мембрану, когда они недостаточно сконцентрированы для самопроизвольной диффузии.Частицы, слишком большие для диффузии или перекачивания, часто проглатываются или выбрасываются целиком при открытии и закрытии мембраны.
Вызывая трансмембранные движения больших молекул, сама клеточная мембрана претерпевает согласованные движения, во время которых часть жидкой среды вне клетки интернализуется (эндоцитоз) или часть внутренней среды клетки выводится наружу (экзоцитоз). Эти движения включают слияние поверхностей мембран с последующим воссозданием неповрежденных мембран.
Эндоцитоз, опосредованный рецепторами Рецепторы играют ключевую роль во многих клеточных процессах. Например, рецептор-опосредованный эндоцитоз позволяет клеткам поглощать молекулы, такие как белки, которые необходимы для нормального функционирования клеток. Encyclopdia Britannica, Inc. Получите эксклюзивный доступ к контенту из нашего 1768 First Edition с подпиской. Подпишитесь сегодня ,нервная система | Определение, функции, структура и факты
Самый простой тип ответа — это прямая индивидуальная реакция на стимул-ответ. Изменение окружающей среды — это стимул; реакция организма на это есть ответ. У одноклеточных организмов реакция является результатом свойства клеточной жидкости, называемого раздражительностью. У простых организмов, таких как водоросли, простейшие и грибы, реакция, при которой организм движется к раздражителю или от него, называется таксисом.У более крупных и сложных организмов — тех, в которых реакция включает синхронизацию и интеграцию событий в различных частях тела, — механизм управления или контроллер расположен между стимулом и реакцией. В многоклеточных организмах этот регулятор состоит из двух основных механизмов, с помощью которых достигается интеграция — химической регуляции и нервной регуляции.
В химической регуляции вещества, называемые гормонами, производятся четко определенными группами клеток и либо диффундируют, либо переносятся кровью в другие области тела, где они действуют на клетки-мишени и влияют на метаболизм или индуцируют синтез других веществ.Изменения, возникающие в результате гормонального действия, выражаются в организме как влияние или изменения в форме, росте, воспроизводстве и поведении.
Растения реагируют на различные внешние раздражители, используя гормоны в качестве регуляторов системы «стимул-ответ». Направленные реакции на движение известны как тропизмы и являются положительными, когда движение направлено к стимулу, и отрицательными, когда оно направлено в сторону от стимула. Когда семя прорастает, растущий стебель поворачивается вверх к свету, а корни поворачиваются вниз, подальше от света.Таким образом, стебель показывает положительный фототропизм и отрицательный геотропизм, тогда как корни демонстрируют отрицательный фототропизм и положительный геотропизм. В этом примере свет и гравитация — это стимулы, а направленный рост — это реакция. Контроллеры — это определенные гормоны, синтезируемые клетками кончиков стеблей растений. Эти гормоны, известные как ауксины, диффундируют через ткани под верхушкой стебля и концентрируются по направлению к затемненной стороне, вызывая удлинение этих клеток и, таким образом, изгиб кончика к свету.Конечным результатом является поддержание растения в оптимальном состоянии с точки зрения освещения.
У животных, помимо химической регуляции через эндокринную систему, существует еще одна интегративная система, называемая нервной системой. Нервную систему можно определить как организованную группу клеток, называемых нейронами, специализирующихся на передаче импульса — возбужденного состояния — от сенсорного рецептора через нервную сеть к эффектору, участку, в котором происходит ответ.
Организмы, обладающие нервной системой, способны к гораздо более сложному поведению, чем организмы, у которых ее нет.Нервная система, специализирующаяся на проведении импульсов, позволяет быстро реагировать на раздражители окружающей среды. Многие реакции, опосредованные нервной системой, направлены на сохранение статус-кво или гомеостаза животного. Стимулы, которые имеют тенденцию перемещать или разрушать какую-либо часть организма, вызывают реакцию, которая приводит к уменьшению неблагоприятных эффектов и возвращению к более нормальному состоянию. Организмы с нервной системой также способны выполнять вторую группу функций, которые инициируют различные модели поведения.Животные могут проходить периоды исследовательского или аппетитного поведения, строительства гнезд и миграции. Хотя эти действия полезны для выживания вида, они не всегда выполняются индивидуумом в ответ на индивидуальную потребность или стимул. Наконец, приобретенное поведение может накладываться как на гомеостатическую, так и на инициирующую функции нервной системы.
Внутриклеточные системы
Все живые клетки обладают свойством раздражительности или отзывчивости на раздражители окружающей среды, которые могут влиять на клетку по-разному, вызывая, например, электрические, химические или механические изменения.Эти изменения выражаются в ответе, который может быть высвобождением секреторных продуктов клетками железы, сокращением мышечных клеток, изгибом стволовой клетки растения или биением ресничных клеток, напоминающих хлыст, «волосков». ,
Отзывчивость отдельной клетки может быть проиллюстрирована поведением относительно простой амебы. В отличие от некоторых других простейших, у амебы отсутствуют высокоразвитые структуры, которые участвуют в приеме стимулов и в производстве или проведении реакции.Однако амеба ведет себя так, как если бы у нее была нервная система, потому что общая отзывчивость ее цитоплазмы служит функциям нервной системы. Возбуждение, производимое стимулом, передается другим частям клетки и вызывает реакцию животного. Амеба переместится в область с определенным уровнем света. Его привлекают химические вещества, выделяемые пищей, и он проявляет реакцию при кормлении. Он также удаляется из области с ядовитыми химическими веществами и проявляет реакцию избегания при контакте с другими объектами.
,Что такое торговля мембранами? | MBInfo
Мембранный трафик включает в себя широкий спектр процессов, которые участвуют в перемещении грузов (обычно белков, патогенов и других макромолекул) с использованием мембраносвязанных транспортных везикул. Этот транспорт может происходить внутри разных органелл в одной и той же клетке или через клеточную мембрану во внеклеточную среду и из нее. Подобно офису сортировки посылок, ячейка использует сложную, строго регулируемую систему, чтобы гарантировать доставку нужного груза в нужное место [1].

Схема, изображающая различные пути мембранного переноса
Мембранный трафик можно разделить на два основных пути в зависимости от направления движения, экзоцитоза и эндоцитоза. Экзоцитоз относится к перемещению груза к плазматической мембране или из клетки. В рамках биосинтетико-секреторного пути вновь синтезированные белки, липиды или углеводы перемещаются из эндоплазматического ретикулума (ER) через Гольджи к клеточной мембране или внеклеточному пространству. И наоборот, эндоцитоз — это перемещение груза в клетку от плазматической мембраны.Это часто можно использовать для поглощения питательных веществ, которые не могут быть синтезированы клеткой, таких как витамины, холестерин и железо. Другая важная функция пути эндоцитоза — направлять грузы на переработку или разложение посредством аутофагии. Клетка также может использовать крупномасштабные эндоцитозные механизмы, такие как фагоцитоз и макропиноцитоз, для интернализации патогенов и внешних частиц с целью поддержания иммунного ответа.
Транспортные везикулы опосредуют перенос через мембрану
И эндоцитоз, и экзоцитоз используют небольшие отсеки мембраны, чтобы заключить свой груз.Эти транспортные пузырьки отпочковываются от одной мембраны и могут динамически сливаться с другими мембранами или расщепляться на более мелкие пузырьки путем деления. Находясь внутри или в просвете транспортных пузырьков, груз защищен от цитоплазмы. Поскольку просвет транспортных везикул физиологически подобен другим мембраносвязанным органеллам и внеклеточному пространству, перенос груза не требует движения через мембрану, а просто слияния везикул.
Транспортные везикулы отличаются друг от друга типом груза, который они переносят с одного места на другое, маршрутом, которым они следуют, а также наличием или отсутствием белков на цитозольной поверхности, которые могут образовывать оболочку.Эти белки оболочки самостоятельно собираются на мембране, помогая собирать и концентрировать груз пузырьков. Существует три хорошо охарактеризованных белка оболочки, которые покрывают везикулы в различных точках эндоцитоза и экзоцитоза. Покрытые клатрином везикулы опосредуют эндоцитоз от плазматической мембраны до эндосомных компартментов и Гольджи. Следующие шаги в эндоцитозе, а именно ретроградный транспорт в пределах Гольджи и к ER, происходит через пузырьки, окруженные белком оболочки I (COPI). В противоположном направлении везикулы COPII отталкиваются от ER секреторным или экзоцитарным путем по направлению к Golgi [2], [3], [4], [5].Кавеолы, которые представляют собой небольшие выпячивания плазматической мембраны в форме луковиц, которые опосредуют эндоцитоз независимо от клатрина, также обладают менее заметной, но окончательной полосатой оболочкой, состоящей из двух белковых комплексов — кавеолинов и кавинов. Были описаны другие клатрин-независимые пути эндоцитов, в которых используются везикулы, лишенные белков оболочки, в основном путь CLIC / GEEC, RhoA-зависимый путь, Arf6-зависимый путь и Flotillin-зависимый путь.
Когда покрытый зачаток везикулы вырастет и будет готов к отсоединению, его необходимо отделить от исходной мембраны без потери груза.В некоторых случаях отслоение отпочковавшихся везикул от плазматической мембраны может быть облегчено с помощью GTPase Dynamin посредством разрыва мембраны. Dynamins связываются на шейке зарождающейся везикулы и сливают два липидных бислоя вместе в GTP-зависимом процессе, тем самым разрезая шейку и высвобождая везикулу из мембраны. Чтобы изгибать мембрану, чтобы способствовать или препятствовать разрыву, белки домена BAR, N-BAR и F-BAR влияют на кривизну мембраны, либо способствуя, либо уменьшая вероятность разрыва мембраны [6], [7].
Цитоскелет отвечает за перемещение пузырьков по клетке. Большинство везикул движется по микротрубочкам с использованием моторов кинезина или динеина, хотя они также могут использовать моторы миозина II и миозина V для перемещения по актиновой сети [8], [9]. Чтобы определить правильное место для доставки груза, транспортная везикула должна распознавать конкретную цель. Члены семейства Rab small GTPase в первую очередь ответственны за обеспечение этой специфичности. Белки Rab могут экспрессироваться как на транспортных везикулах, так и на мембранах-мишенях, обеспечивая дополнительный уровень регуляции.Белки SNARE стыковывают транспортную везикулу в правильном месте на мембране и катализируют слияние мембран, заключительный этап доставки груза. SNARE сближают соприкасающиеся мембраны транспортной везикулы и целевой области, так что липиды из разных бислоев могут смешиваться, и это в конечном итоге приводит к слиянию и высвобождению груза [10], [7].
,