БИОЛОГИЧЕСКИЕ МЕМБРАНЫ • Большая российская энциклопедия
БИОЛОГИ́ЧЕСКИЕ МЕМБРА́НЫ (лат. membrana – кожица, оболочка, перепонка), структуры, ограничивающие содержимое клеток (клеточная, или плазматическая, мембрана, плазмалемма) и внутриклеточных органелл. У прокариот имеется только клеточная мембрана, в большинстве случаев окружённая клеточной стенкой. У эукариот мембраной окружена не только клетка, но и ядро, а также митохондрии, лизосомы, пероксисомы, секреторные гранулы, эндосомы, у растений ещё – хлоропласты и вакуоли; мембраны образуют также разветвлённую сеть эндоплазматического ретикулума и комплекса Гольджи. Митохондрии, хлоропласты и ядра окружены двумя мембранами, а внутри хлоропластов имеется ещё один тип мембран, формирующих тилакоиды. У животных к клеточной мембране снаружи примыкает гликопротеиновый комплекс – гликокаликс, у растений – клеточная стенка. Толщина мембран варьирует от 6 до 10 нм.
Структура биологических мембран
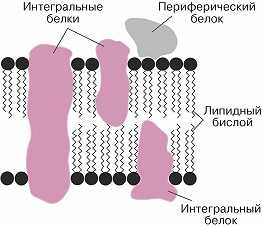
Схема строения клеточной мембраны.
Основу Б. м. составляет протяжённый двойной слой (бислой) глицерофосфо-, сфинго- и гликолипидов со встроенными в него молекулами различных белков. Гидрофобные (неполярные) группы молекул липидов (остатки жирных кислот) погружены в толщу мембраны, а гидрофильные (полярные) головки ориентированы наружу, в окружающую водную среду (см. Липиды). Плотность упаковки Б. м. обеспечивается электростатическими взаимодействиями полярных головок и гидрофобными контактами между цепями жирных кислот. Входящие в состав Б. м. белки взаимодействуют с липидным бислоем с помощью гидрофобных взаимодействий и вандерваальсовых связей. Соотношение липидов и белков, их состав в разл. Б. м. могут существенно различаться. Так, в мембранах миелиновой оболочки содержание липидов (по массе) в четыре раза больше, чем белков, а во внутр. мембранах митохондрий более чем в два раза преобладают белки. Липиды Б. м. представлены гл. обр. фосфатидилхолином, фосфатидилэтаноламином, сфингомиелином, фосфатидилсерином, фосфатидилинозитом и кардиолипином, которые обнаруживаются примерно в одном и том же соотношении в мембранах разных по уровню организации организмов. В то же время набор жирных кислот, входящих в состав липидов, подвержен изменениям. Напр., понижение темп-ры, давления и солёности среды обитания организмов сопровождаются увеличением количества ненасыщенных связей и/или короткоцепочечных жирных кислот в фосфолипидах и происходящим вследствие этого уменьшением плотности упаковки бислоя. Липидам свойственна определённая подвижность внутри бислоя. Они способны к быстрому вращению вокруг оси (вращательная диффузия), к свободному перемещению в пределах одного слоя мембраны (латеральная диффузия), а также к переходу с одной стороны бислоя на другую (такое передвижение обеспечивается спец. механизмами). Для клеточных мембран животных клеток характерно высокое содержание холестерина (в ср. ок. 21%), который участвует в регуляции текучести мембраны, препятствуя плотной упаковке фосфолипидов. В растит. клетке роль холестерина играет его аналог – десмостерин. В мембранах бактерий и внутриклеточных органелл стерины отсутствуют. До 10% сухого вещества мембран приходится на долю углеводов, которые экспонированы на внешней стороне клеточной мембраны и являются составной частью мембранных гликолипидов и гликопротеинов.
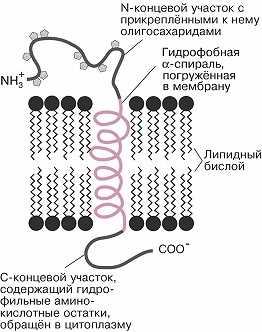
Схема структурной организации интегрального белка. Спиральный участок встроен в гидрофобную часть липидного бислоя; N-концевой участок с прикреплёнными к нему олигосахаридными цепями расположен на вне…
Содержание белка в разл. мембранах колеблется от 20 до 75% (в пересчёте на сухую массу). Мембранные белки могут быть встроены в бислой (интегральные белки). При этом они погружены в мембрану и пронизывают её (иногда неск. раз) таким образом, что достаточно протяжённые участки белка, образованные гидрофобными аминокислотами, оказываются в её толще, а гидрофильные – на поверхности, по обе стороны Б. м. Выступающие над внешней стороной мембраны участки белковых молекул обычно несут неск. ковалентно связанных, часто разветвлённых цепей олигосахаридов, образованных остатками маннозы, фукозы, глюкозы, N-ацетилглюкозамина и др. Эти компоненты играют роль маркеров при распознавании клеточной поверхности. Молекулы периферических белков расположены гл. обр. на внутренней поверхности мембраны, не проникая внутрь бислоя, и удерживаются на ней с помощью электростатич. взаимодействий и водородных связей; они связываются с мембраной обратимо и могут переходить в цитоплазму при модификации белков (напр., путём их фосфорилирования) в ответ на изменения функционального состояния клетки. Мн. белки организованы в виде сложных комплексов (напр., белки дыхательной цепи митохондрий). В клетках происходит постоянное обновление компонентов Б. м. путём введения новых молекул липидов и белков, однако структурная организация Б. м. в течение всей жизни клетки остаётся неизменной.
Функции мембран
Осн. функции мембран связаны с белками. Мн. мембранные белки – ферменты, обеспечивающие протекание окислительно-восстановительных, гидролитич. и биосинтетич. реакций как на поверхности мембраны, так и внутри неё. Важнейшая функция белков мембран – транспортная. Жирорастворимые соединения (напр., стероидные гормоны) легко проникают в липидный бислой, но для большинства др. соединений (в т. ч. аминокислот, сахаров) и неорганич. ионов он непроницаем. Специально предназначенные для этих целей мембранные белки обеспечивают как активный (требующий затрат энергии), так и пассивный (за счёт градиента концентраций) транспорт веществ и ионов (см. Ионные каналы). Специфич. белки – ионные насосы ($\ce{Na/K}$-насос и $\ce{Ca}$-насос) ответственны за асимметричное распределение ионов $\ce{Na^+, K^+}$ и $\ce{Ca^{2+}}$ по обе стороны клеточной мембраны (напр., в цитоплазме животной клетки – низкую для $\ce{Na^+}$ и $\ce{Ca^{2+}}$ и высокую для $\ce{K^+}$). Такая асимметрия обеспечивает мн. проявления жизнедеятельности (электровозбудимость, осмотическую устойчивость и др.). Мембранные белки аквапорины образуют в мембране спец. каналы, регулирующие проникновение в клетку молекул воды. Взаимодействие клетки с внешней средой, регуляция внутриклеточных процессов осуществляются посредством рецепторных белков (рецепторов), ответственных за фото-, термо-, механо- и хеморецепцию.
Барьерная функция Б. м. обеспечивает сохранение определённого состава клетки и концентрации составляющих её веществ, а также защиту от воздействия разл. чужеродных факторов и токсинов. Благодаря Б. м. внутри клеток возможно одноврем. протекание множества несовместимых друг с другом реакций. Напр., необходимые клетке белки синтезируются на рибосомах, прикреплённых к эндоплазматическому ретикулуму, а их распад происходит в лизосомах. В Б. м. протекают процессы энергообмена клеток. Внутр. мембраны митохондрий и мембраны тилакоидов – важнейшие преобразователи энергии, играющие ключевую роль в запасании энергии, образующейся в ходе дыхания и фотосинтеза, в энергию пирофосфатной связи аденозинтрифосфата. Б. м. нейронов могут генерировать и осуществлять передачу электрич. сигнала, участвуя тем самым в процессах возбуждения и проведения нервного импульса.
Белковые и липидные компоненты выполняют ряд др. функций. Фрагменты фосфолипидов могут выступать в качестве предшественников сигнальных молекул (мессенджеров). Напр., при активации мембранной фосфолипазы А из бислоя высвобождается арахидоновая кислота, дальнейшие превращения которой приводят к образованию биологич. регуляторов – тромбоксанов, лейкотриенов и простагландинов. Фосфатидилсерин, локализованный на внутр. стороне мембраны, при инициации апоптоза мигрирует на её внешнюю сторону. Его появление служит сигналом для фагоцитов, которые имеют рецепторы на этот фосфолипид; они «узнают» дефектные клетки и уничтожают их. Гликолипиды наряду с гликопротеинами играют важную роль в явлениях межклеточной адгезии, участвуют в иммунных реакциях.
Методы изучения биологических мембран
Для изучения структуры и функции мембран используются электрофизиологич. и иммуноцитохимич. методы, жидкостная хроматография (для идентификации и анализа липидных компонентов), проточная цитометрия, позволяющая проследить ответ клетки на взаимодействие специфич. лигандов с клеточной мембраной, разнообразные физич. методы, характеризующие структуру мембран, упаковку и подвижность липидов в бислое (в т. ч. электронная микроскопия, малоугловое рассеивание нейтронов, флуоресцентная спектроскопия, круговой дихроизм), и др. методы.
Разнообразие типов Б. м., их полифункциональность и высокая чувствительность к внешним воздействиям являются причиной того, что они вовлекаются в разл. патологич. процессы. Повреждения клеточных мембран, приводящие к образованию свободных радикалов и гибели нервных клеток, лежат в основе нейродегенеративных заболеваний (болезнь Альцгеймера, паркинсонизм, боковой амиотрофический склероз), могут служить причиной инсульта и инфаркта миокарда.
bigenc.ru
11.Био мембраны, их строение и функции.
Био мембраны-функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей. Биологические мембраны имеются во всех клетках. Их значение определяется важностью функций, которые они выполняют в процессе нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при различных нарушениях мембранных функций и проявляющихся практически на всех уровнях организации — от клетки и субклеточных систем до тканей, органов и организма в целом. Мембранные структуры клетки представлены поверхностной (клеточной, или плазматической) и внутриклеточными (субклеточными) мембранами. Название внутриклеточных (субклеточных) мембран обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и др. (см. Клетка). Толщина биологических мембран — 7—10 нм, но их общая площадь очень велика, например, в печени крысы она составляет несколько сот квадратных метров.
Химический состав и строение биологических мембран. Состав М.б. зависит от их типа и функций, однако основными составляющими являются липиды и белки, а также углеводы(небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса). Липиды. В составе М.б. обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды. В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит) и сфингофосфолипиды (производные церамида, сфингомиелин). Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%). В липидных компонентах М.б. содержатся разнообразные жирные кислоты, однако в мембранах животных клеток преобладают пальмитиновая, олеиновая и стеариновая кислоты. Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать двухслойные структуры (бислои) при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» —наоборот, наружу. Липидный бислои представляет собой динамичную структуру: образующие его липиды могут вращаться, двигаться в латеральном направлении и даже переходить из слоя в слой (флип-флоп переход). Такое строение липидного бислоя легло в основу современных представлений о структуре М.б. и определяет некоторые важные свойства М.б., например способность служить барьером и не пропускать молекулы веществ, растворенных в воде Нарушение структуры бислоя может привести к нарушению барьерной функции мембран. Холестерин в составе М.б. играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов. Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность. Белки биологических мембран исключительно разнообразны. Молекулярная масса их в большинстве своем составляет 25 000 — 230 000. Белки могут взаимодействовать с липидным бислоем за счет электростатических и (или) межмолекулярных сил. Они сравнительно легко могут быть удалены из мембраны. К такому типу белков относят цитохром с (молекулярная масса около 13 000), обнаруживаемый на наружной поверхности внутренней мембраны митохондрий. Эти белки называются периферическими, или наружными. Для других белков, получивших название интегральных, или внутренних, характерно то, что одна или несколько полипептидных цепей оказываются погруженными в бислои или пересекают его, иногда не один раз (например, гликофорин, транспортные АТФ-азы, бактериородопсин). Часть белка, контактирующая с гидрофобной частью липидного бислоя, имеет спиральное строение и состоит из неполярных аминокислот, в силу чего между этими компонентами белков и липидов происходит гидрофобное взаимодействие. Полярные группы гидрофильных аминокислот непосредственно взаимодействуют с примембранными слоями, как с одной, так и с другой стороны бислоя. Молекулы белков, как и молекулы липидов, находятся в динамическом состоянии, для них также характерна вращательная, латеральная и вертикальная подвижность. Она является отражением не только их собственной структуры, но и функциональной активности. что в значительной степени определяется вязкостью липидного бислоя, которая, в свою очередь, зависит от состава липидов, относительного содержания и вида ненасыщенных жирно-кислотных цепей. Этим объясняется узкий температурный диапазон функциональной активности мембраносвязанных белков. Белки мембран выполняют три основные функции: каталитическую (ферменты), рецепторную и структурную. Однако такое разграничение достаточно условно, и в ряде случаев один и тот же белок может выполнять и репепторную и ферментную функции (например, инсулин). Число
Функции биологических мембран. Барьерная функция. Для клеток и субклеточных частиц М.б. служат механическим барьером, отделяющим их от внешнего пространства. Функционирование клетки часто сопряжено с наличием значительных механических градиентов на ее поверхности преимущественно вследствие осмотического и гидростатического давления. Основную нагрузку в этом случае несет клеточная стенка, главными структурными элементами которой у высших растений являются целлюлоза, пектин и экстепин, а у бактерий — муреин (сложный полисахарид-пептид). В клетках животных необходимость в жесткой оболочке отсутствует. Некоторую жесткость этим клеткам придают особые белковые структуры цитоплазмы, примыкающие к внутренней поверхности плазматической мембраны. Перенос веществ через М.б. сопряжен с такими важнейшими биологическими явлениями, как внутриклеточный гомеостаз ионов, биоэлектрические потенциалы, возбуждение и проведение нервного импульса, запасание и трансформация энергии и т.п. (см. Биоэнергетика). Различают пассивный и активный транспорт (перенос) нейтральных молекул, воды и ионов через М.б. Пассивный транспорт не связан с затратами энергии, он осуществляется путем диффузии по концентрационным, электрическим или гидростатическим градиентам. Активный транспорт осуществляется против градиентов, связан с затратой энергии (преимущественно энергии гидролиза АТФ) и сопряжен с работой специализированных мембранных систем (мембранных насосов). Различают несколько видов транспорта. Если вещество транспортируется через мембрану независимо от наличия и переноса других соединений, то такой вид транспорта называют юнипортом. Если перенос одного вещества сопряжен с транспортом другого, то говорят о котранспорте, причем однонаправленный перенос называется симпортом, а противоположно направленный — антипортом. В особую группу выделяют перенос веществ путем экзо- и пиноцитоза. Пассивный перенос может осуществляться путем простой диффузии через липидный бислои мембраны, а также через специализированные образования — каналы. Путем диффузии через мембрану проникают в клетку незаряженные молекулы, хорошо растворимые в липидах, в т.ч. многие яды и лекарственные средства, а также кислород и углекислый газ. Каналы представляют собой липопротеиновые структуры, пронизывающие мембраны. Они служат для переноса определенных ионов и могут находиться в открытом или закрытом состоянии. Проводимость канала зависит от мембранного потенциала, что играет важную роль в механизме генерации и проведения нервного импульса. В ряде случаев перенос вещества совпадает с направлением градиента, но существенно превосходит по скорости простую диффузию. Этот процесс называют облегченной диффузией; он происходит с участием белков-переносчиков. Процесс облегченной диффузии не нуждается в энергии. Этим способом транспортируются сахара, аминокислоты, азотистые основания. Такой процесс происходит, например, при всасывании сахаров из просвета кишечника клетками эпителия. Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 106, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума — 104, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более 1/3 всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия — натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса — Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ. На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая — аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида. Изложенные механизмы транспорта различных веществ через клеточные мембраны имеют место и в случае их транспорта через эпителий ряда органов (кишечника, почек, легких), который осуществляется через слой клеток (монослой в кишечнике и нефронах), а не через единичную клеточную мембрану. Такой транспорт называют трансцеллюлярным, или трансэпителиальным. Характерной особенностью клеток, например эпителиоцитов кишечника и канальцев нефронов, является то, что апикальная и базальная их мембраны различаются по проницаемости, величине мембранного потенциала и транспортной функции. Способность генерировать биоэлектрические потенциалы и проводить возбуждение. Возникновение биоэлектрических потенциалов связано с особенностями строения биологических мембран и с деятельностью их транспортных систем, создающих неравномерное распределение ионов по обе стороны мембраны (см. Биоэлектрические потенциалы,Возбуждение). Процессы трансформации и запасания энергии протекают в специализированных М.б. и занимают центральное место в энергетическом обеспечении живых систем. Два основных процесса энергообразования — фотосинтез и тканевое дыхание — локализованы в мембранах внутриклеточных органелл высших организмов, а у бактерий — в клеточной (плазматической) мембране (см. Дыхание тканевое). Фотосинтезирующие мембраны преобразуют энергию света в энергию химических соединений, запасая ее в форме сахаров — основного химического источника энергии для гетеротрофных организмов. При дыхании энергия органических субстратов освобождается в процессе переноса электронов по цепи окислительно-восстановительных переносчиков и утилизируется в процессе фосфорилирования АДФ неорганическим фосфатом с образованием АТФ. Мембраны, осуществляющие фосфорилирование, сопряженное с дыханием, называют сопрягающими (внутренние мембраны митохондрий, клеточные мембраны некоторых аэробных бактерий, мембраны хроматофоров фотосинтезирующих бактерий). Метаболические функции мембран определяются двумя факторами: во-первых, связью большого числа ферментов и ферментативных систем с мембранами, во-вторых, способностью мембран физически разделять клетку на отдельные отсеки, отграничивая друг от друга метаболические процессы, протекающие в них. Метаболические системы не остаются при этом полностью изолированными. В мембранах, разделяющих клетку, имеются специальные системы, обеспечивающие избирательное поступление субстратов, выделение продуктов, а также движение соединений, обладающих регуляторным действием. Клеточная рецепция и межклеточные взаимодействия. Под этой формулировкой объединен весьма обширный и разнообразный набор важных функций клеточных мембран, определяющих взаимодействие клетки с окружающей средой и формирование многоклеточного организма как единого целого. Молекулярно-мембранные аспекты клеточной рецепции и межклеточных взаимодействий касаются прежде всего иммунных реакций, гормонального контроля роста и метаболизма, закономерностей эмбрионального развития.
studfiles.net
Биологические мембраны
Биологические мембраны
Содержание
Введение
. Мембраны биологические
. Химический состав и строение биологических мембран
.1 Липиды биологических мембран
.2 Белки биологических мембран
.3 Углеводы биологических мембран
. Свойства (функции) биологических мембран
.1 Барьерная функция
.2 Перенос веществ
3.3 Способность генерировать биоэлектрические потенциалы и проводить возбуждение
3.4 Процессы трансформации и запасания энергии
.5 Метаболические свойства мембран
.6 Клеточная рецепция и межклеточные взаимодействия
. Нарушения структуры и функций биологических мембран
Заключение
Список использованных источников
Введение
Мембраны биологические (лат. membrana оболочка, перепонка) — это функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей.
Биологические мембраны имеются во всех клетках. Их значение определяется важностью функций, которые они выполняют в процессе нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при различных нарушениях мембранных функций и проявляющихся практически на всех уровнях организации — от клетки и субклеточных систем до тканей, органов и организма в целом. Вышесказанное определяет актуальность работы.
В работе использованы научные публикации Болдырева А.А., Конева С.В., Мажуля В.М., Кульберга А.Я., Маленкова А.Г., Сима Э., Финеана Дж., Колмэна Р., Митчелл Р. и других.
Структурно работа состоит из введения, 4 глав, заключения и списка использованных источников.
1. Мембраны биологические
Мембранные структуры клетки представлены поверхностной (клеточной, или плазматической) и внутриклеточными (субклеточными) мембранами. Название внутриклеточных (субклеточных) мембран обычно зависит от названия ограничиваемых или образуемых ими структур. Так, различают митохондриальные, ядерные, лизосомные мембраны, мембраны пластинчатого комплекса аппарата Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и др. Толщина биологических мембран — 7-10 нм, но их общая площадь очень велика, например, в печени крысы она составляет несколько сот квадратных метров.
Схема строения биологической мембраны изображена на рисунке 1.
Термин «мембраны» как окружающей клетку невидимой плёнки, служащей барьером между содержимым клетки и внешней средой и одновременно — полупроницаемой перегородкой, через которую могут проходить вода и некоторые растворенные в ней вещества, был впервые использован, ботаниками фон Молем и независимо К. фон Негели в 1855 г для объясненеия явлений плазмолиза. В 1877 г. ботаник В. Пфеффер (1845-1920) опубликовал свой труд «Исследования осмоса» (Leipzig), где описал существование клеточных мембран.
В 1890 году немецкий физико-химик и философ В. Оствальд обратил внимание на возможную роль мембран в биоэлектрических процессах. Между 1895 и 1902 годами Э. Овертон измерил проницаемость клеточной мембраны для большого числа соединений и показал прямую зависимость между способностью этих соединений проникать через мембраны и их растворимостью в липидах. Это было чётким указанием на то, что именно липиды формируют плёнку, через которую проходят в клетку вещества из окружающего раствора.
Рис. 1. Схема строения биологической мембраны: 1 — углеводные фрагменты гликопротеидов; 2 — липидный бислой; 3 — интегральный белок; 4 — «головки» фосфолипидов; 5 — периферический белок; 6 — холестерин; 7 — жирнокислотные «хвосты» фосфолипидов.
В 1925 году Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. На основе результатов этих исследований было сделано предположение, что липиды в мембране располагаются в виде бимолекулярного слоя. Вместе с тем имелись экспериментальные данные, которые свидетельствовали о том, что биологическая мембрана содержит в своем составе и белковые молекулы. Эти противоречия экспериментальных результатов были устранены Даниелли и Давсоном, предложившими в 1935 году так сказать «бутербродную» модель строения биологических мембран, которая с некоторыми несущественными изменениями продержалась в мембранологии в течении почти 40 лет. Согласно этой модели, на поверхности фосфолипидного бислоя в мембранах располагаются белки.
2. Химический состав и строение биологических мембран
Состав биологических мембран зависит от их типа и функций, однако основными составляющими являются липиды и белки, а также углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
.1 Липиды биологических мембран
В составе биологических мембран обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды.
В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды и сфингофосфолипиды. Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%).
В липидных компонентах биологических мембран содержатся разнообразные жирные кислоты, однако в мембранах животных клеток преобладают пальмитиновая, олеиновая и стеариновая кислоты.
Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать двухслойные структуры (бислои) при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» -наоборот, наружу. Липидный бислои представляет собой динамичную структуру: образующие его липиды могут вращаться, двигаться в латеральном направлении и даже переходить из слоя в слой (флип-флоп переход). Такое строение липидного бислоя легло в основу современных представлений о структуре биологических мембран и определяет их некоторые важные свойства, например способность служить барьером и не пропускать молекулы веществ, растворенных в воде. Нарушение структуры бислоя может привести к нарушению барьерной функции мембран.
Холестерин в составе биологических мембран играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов.
Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность.
.2 Белки биологических мембран
Белки биологических мембран исключительно разнообразны.
Белки могут взаимодействовать с липидным бислоем за счет электростатических и (или) межмолекулярных сил. Они сравнительно легко могут быть удалены из мембраны. К такому типу белков относят цитохром, обнаруживаемый на наружной поверхности внутренней мембраны митохондрий. Эти белки называются периферическими, или наружными. Для других белков, получивших название интегральных, или внутренних, характерно то, что одна или несколько полипептидных цепей оказываются погруженными в бислои или пересекают его, иногда не один раз (например, гликофорин, транспортные АТФ-азы, бактериородопсин).
Часть белка, контактирующая с гидрофобной частью липидного бислоя, имеет спиральное строение и состоит из неполярных аминокислот, в силу чего между этими компонентами белков и липидов происходит гидрофобное взаимодействие. Полярные группы гидрофильных аминокислот непосредственно взаимодействуют с примембранными слоями, как с одной, так и с другой стороны бислоя. Молекулы белков, как и молекулы липидов, находятся в динамическом состоянии, для них также характерна вращательная, латеральная и вертикальная подвижность. Она является отражением не только их собственной структуры, но и функциональной активности. что в значительной степени определяется вязкостью липидного бислоя, которая, в свою очередь, зависит от состава липидов, относительного содержания и вида ненасыщенных жирно-кислотных цепей. Этим объясняется узкий температурный диапазон функциональной активности мембраносвязанных белков.
Белки мембран выполняют три основные функции: каталитическую (ферменты), рецепторную и структурную. Однако такое разграничение достаточно условно, и в ряде случаев один и тот же белок может выполнять и репепторную и ферментную функции (например, инсулин).
Число мембранных ферментов в клетке достаточно велико, однако их распределение в различных типах биологических мембран неодинаково. Некоторые ферменты (маркерные) присутствуют только в мембранах определенного типа. Ннапример, аденилатциклаза — в плазматической мембране; НАДФН-дегидрогеназа, цитохром в 5 — в мембранах эндоплазматического ретикулума; моноаминоксидаза — в наружной мембране митохондрий, а сукцинат-дегидрогеназа — во внутренней; кислая фосфатаза — в мембране лизосом.
Рецепторные белки, специфически связывая низкомолекулярные вещества (многие гормоны, медиаторы), обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка принимает различные сигналы, поступающие из внешней среды.
К структурным белкам относят белки цитоскелета, прилегающие к цитоплазматической стороне клеточной мембраны. В комплексе с микротрубочками и микрофиламентами цитоскелета они обеспечивают противодействие клетки изменению ее объема и создают эластичность. В эту же группу включают ряд мембранных белков, функции которых не установлены.
.3 Углеводы биологических мембран
Углеводы в биологических мембранах находятся в соединении с белками (гликопротеины) и липидами (гликолипиды). Углеводные цепи белков представляют собой олиго- или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Углеводные компоненты биологических мембран открываются в основном во внеклеточную среду, образуя на поверхности клеточных мембран множество ветвистых образований, являющихся фрагментами гликолипидов или гликопротеидов. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обеспечением стабильности белковых молекул в биологической мембране. Многие рецепторные белки содержат углеводные компоненты. Примером могут служить антигенные детерминанты групп крови, представленные гликолипидами и гликопротеинами.
3. Свойства (функции) биологических мембран
.1 Барьерная функция
Для клеток и субклеточных частиц биологические мембран служат механическим барьером, отделяющим их от внешнего пространства. Функционирование клетки часто сопряжено с наличием значительных механических градиентов на ее поверхности преимущественно вследствие осмотического и гидростатического давления. Основную нагрузку в этом случае несет клеточная стенка, главными структурными элементами которой у высших растений являются целлюлоза, пектин и экстепин, а у бактерий — муреин (сложный полисахарид-пептид). В клетках животных необходимость в жесткой оболочке отсутствует. Некоторую жесткость этим клеткам придают особые белковые структуры цитоплазмы, примыкающие к внутренней поверхности плазматической мембраны.
.2 Перенос веществ
Перенос веществ через биологические мембраны сопряжен с такими важнейшими биологическими явлениями, как внутриклеточный гомеостаз ионов, биоэлектрические потенциалы, возбуждение и проведение нервного импульса, запасание и трансформация энергии и т.п.
.Различают пассивный и активный транспорт (перенос) нейтральных молекул, воды и ионов через биологические мембраны.
Пассивный транспорт не связан с затратами энергии, он осуществляется путем диффузии по концентрационным, электрическим или гидростатическим градиентам (разностью показателей по обеим сторонам мембраны).
Активный транспорт осуществляется против градиентов, связан с затратой энергии (преимущественно энергии гидролиза АТФ) и сопряжен с работой специализированных мембранных систем (мембранных насосов). Различают несколько видов транспорта. Если вещество транспортируется через мембрану независимо от наличия и переноса других соединений, то такой вид транспорта называют юнипортом. Если перенос одного вещества сопряжен с транспортом другого, то говорят о котранспорте, причем однонаправленный перенос называется симпортом, а противоположно направленный — антипортом. В особую группу выделяют перенос веществ путем экзо- и пиноцитоза.
Пассивный перенос может осуществляться путем простой диффузии через липидный бислои мембраны, а также через специализированные образования — каналы. Путем диффузии через мембрану проникают в клетку незаряженные молекулы, хорошо растворимые в липидах, в т.ч. многие яды и лекарственные средства, а также кислород и углекислый газ. Каналы представляют собой липопротеиновые структуры, пронизывающие мембраны. Они служат для переноса определенных ионов и могут находиться в открытом или закрытом состоянии. Проводимость канала зависит от мембранного потенциала, что играет важную роль в механизме генерации и проведения нервного импульса.
В ряде случаев перенос вещества совпадает с направлением градиента, но существенно превосходит по скорости простую диффузию.
Этот процесс называют облегченной диффузией; он происходит с участием белков-переносчиков. Процесс облегченной диффузии не нуждается в энергии. Этим способом транспортируются сахара, аминокислоты, азотистые основания. Такой процесс происходит, например, при всасывании сахаров из просвета кишечника клетками эпителия.
Перенос молекул и ионов против электрохимического градиента (активный транспорт) связан со значительными затратами энергии. Часто градиенты достигают больших величин. например, концентрационный градиент водородных ионов на плазматической мембране клеток слизистой оболочки желудка составляет 106, градиент концентрации ионов кальция на мембране саркоплазматического ретикулума — 104, при этом потоки ионов против градиента значительны. В результате затраты энергии на транспортные процессы достигают, например, у человека, более трети всей энергии метаболизма. В плазматических мембранах клеток различных органов обнаружены системы активного транспорта ионов натрия и калия — натриевый насос. Эта система перекачивает натрий из клетки и калий в клетку (антипорт) против их электрохимических градиентов. Перенос ионов осуществляется основным компонентом натриевого насоса — Na+, К+-зависимой АТФ-азой за счет гидролиза АТФ (аденозинтрифосфата).
На каждую гидролизующуюся молекулу АТФ транспортируется три иона натрия и два иона калия. Существуют два типа Са2+-АТФ-аз. Одна из них обеспечивает выброс ионов кальция из клетки в межклеточную среду, другая — аккумуляцию кальция из клеточного содержимого во внутриклеточное депо. Обе системы способны создавать значительный градиент иона кальция. К+, Н+-АТФ-аза обнаружена в слизистой оболочке желудка и кишечника. Она способна транспортировать Н+ через мембрану везикул слизистой оболочки при гидролизе АТФ. В микросомах слизистой оболочки желудка лягушки найдена аниончувствительная АТФ-аза, способная при гидролизе АТФ осуществлять антипорт бикарбоната и хлорида.
Изложенные механизмы транспорта различных веществ через клеточные мембраны имеют место и в случае их транспорта через эпителий ряда органов (кишечника, почек, легких), который осуществляется через слой клеток (монослой в кишечнике и нефронах), а не через единичную клеточную мембрану. Такой транспорт называют трансцеллюлярным, или трансэпителиальным. Характерной особенностью клеток, например эпителиоцитов кишечника и канальцев нефронов, является то, что апикальная и базальная их мембраны различаются по проницаемости, величине мембранного потенциала и транспортной функции.
3.3 Способность генерировать биоэлектрические потенциалы и проводить возбуждение
Возникновение биоэлектрических потенциалов связано с особенностями строения биологических мембран и с деятельностью их транспортных систем, создающих неравномерное распределение ионов по обе стороны мембраны.
Внутри клетки, находящейся в состоянии покоя, содержится ионов калия в 30 раз больше, чем во внеклеточной жидкости, натрия же во внеклеточной жидкости содержится в 20 раз больше, чем в клетке.
В соответствии с градиентом концентрации калий стремится выйти из клетки, натрий — войти в клетку. Полупроницаемая мембрана пропускает ионы калия, которые выходят из клетки и располагаются на внешней поверхности мембраны. На внутренней поверхности мембраны сосредоточиваются отрицательно заряженные ионы.
При возбуждении (деполяризации) мембрана становится проницаемой для ионов натрия, который устремляется внутрь клетки; внутренняя поверхность мембраны заряжается положительно, внешняя — отрицательно.
Накопление положительных ионов внутри клетки и освободившихся отрицательных ионов снаружи приведет к «обратной поляризации», наружная поверхность мембраны приобретает отрицательный заряд, внутренняя — положительный. Затем поток натрия внутрь клетки уменьшается, так как понижается проницаемость мембраны для натрия, выход ионов калия из клетки начинает превышать ток ионов натрия в клетку, наступает период реполяризации (восстановление исходной поляризации).
.4 Процессы трансформации и запасания энергии
мембрана клеточный биоэлектрический фотосинтез
Процессы трансформации и запасания энергии протекают в специализированных биологических мембранах и занимают центральное место в энергетическом обеспечении живых систем. Два основных процесса энергообразования — фотосинтез и тканевое дыхание — локализованы в мембранах внутриклеточных органелл высших организмов, а у бактерий — в клеточной (плазматической) мембране.
Фотосинтезирующие мембраны преобразуют энергию света в энергию химических соединений, запасая ее в форме сахаров — основного химического источника энергии для гетеротрофных организмов. При дыхании энергия органических субстратов освобождается в процессе переноса электронов по цепи окислительно-восстановительных переносчиков и утилизируется в процессе фосфорилирования АДФнеорганическим фосфатом с образованием АТФ.
Мембраны, осуществляющие фосфорилирование, сопряженное с дыханием, называют сопрягающими (внутренние мембраны митохондрий, клеточные мембраны некоторых аэробных бактерий, мембраны хроматофоров фотосинтезирующих бактерий).
.5 Метаболические свойства мембран
Метаболические функции мембран определяются двумя факторами:
во-первых, связью большого числа ферментов и ферментативных систем с мембранами;
во-вторых, способностью мембран физически разделять клетку на отдельные отсеки, отграничивая друг от друга метаболические процессы, протекающие в них.
Метаболические системы не остаются при этом полностью изолированными. В мембранах, разделяющих клетку, имеются специальные системы, обеспечивающие избирательное поступление субстратов, выделение продуктов, а также движение соединений, обладающих регуляторным действием.
3.6 Клеточная рецепция и межклеточные взаимодействия
Под этой формулировкой объединен весьма обширный и разнообразный набор важных функций клеточных мембран, определяющих взаимодействие клетки с окружающей средой и формирование многоклеточного организма как единого целого.
Под клеточной рецепцией понимают процесс восприятия и преобразования химического сигнала в сложную последовательность внутриклеточных химических процессов. Клеточная рецепция обеспечивает возможность обмена информации между клетками, который осуществляется при помощи биологически активных веществ (гормонов, медиаторов). Обязательным этапом такого межклеточного взаимодействия является связывание молекул вещества с соответствующей молекулой клетки-мишени, называемой клеточным рецептором. Роль клеточных рецепторов играют специфические белковые молекулы, которые могут быть расположены на поверхности клетки, в цитоплазме или в ядре.
Молекулярно-мембранные аспекты клеточной рецепции и межклеточных взаимодействий касаются прежде всего иммунных реакций, гормонального контроля роста и метаболизма, закономерностей эмбрионального развития.
4. Нарушения структуры и функции биологических мембран
Разнообразие типов биологических мембран, их полифункциональность и высокая чувствительность к внешним условиям порождают необыкновенное разнообразие структурно-функциональных нарушений мембран, возникающих при многих неблагоприятных воздействиях и сопряженных с огромным числом конкретных заболеваний организма как целого. Все это разнообразие нарушений достаточно условно можно подразделить на:
транспортные;
функционально-метаболические;
структурные.
В общем виде охарактеризовать последовательность возникновения этих нарушений не представляется возможным, и в каждом конкретном случае требуется детальный анализ для выяснения первичного звена в цепи развития структурно-функциональных нарушений мембран. Нарушение транспортных функций мембран, в частности увеличение проницаемости мембран, — общеизвестный универсальный признак повреждения клетки.
Нарушением транспортных функций (например, у человека) обусловлено более 20 так называемых транспортных болезней, среди которых почечная глюкозурия, цистинурия, нарушение всасывания глюкозы, галактозы и витамина В12, наследственный сфероцитоз и др.
Среди функционально-метаболических нарушений биологических мембран центральными являются изменения процессов биосинтеза, а также многообразные отклонения в энергообеспечении живых систем. В наиболее общем виде следствием этих процессов является нарушение состава и физико-химических свойств мембран, выпадение отдельных звеньев метаболизма и его извращение, а также снижение уровня жизненно важных энергозависимых процессов (активного транспорта ионов, процессов сопряженного транспорта, функционирования сократительных систем и т.д.). Повреждения ультраструктурной организации биологических мембран выражаются в чрезмерном везикулообразовании, увеличении поверхности плазматических мембран за счет образования пузырей и отростков, слиянии разнородных клеточных мембран, образовании микропор и локальных структурных дефектов.
Заключение
В результате выполненной работы сделаны следующие выводы:
. Мембраны биологические — это функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки, они образуют единую внутриклеточную систему канальцев, складок, замкнутых областей.
. В составе биологических мембран основными составляющими являются липиды и белки, присутствуют углеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
. Биологические мембраны осуществляют барьерная функция, перенос веществ, трансформируют и запасают энергию. Они способны генерировать биоэлектрические потенциалы и проводить возбуждение, осуществлять клеточную рецепцию и межклеточные взаимодействия, участвуют в метаболизме клетки.
. Структурно-функциональные нарушения мембран, возникающих при многих неблагоприятных воздействиях приводят к большому количеству заболеваний как отдельных частей (органов) организма, так и всего организма в целом.
Список использованных источников
1.Биологические мембраны, под ред. Д.С. Парсонса, пер. с англ., М., 2008.
2.Болдырев А.А. Введение в биохимию мембран, М., 2006,
.Конев С.В. и Мажуль В.М. Межклеточные контакты. Минск, 2007.
.Кульберг А.Я. Рецепторы клеточных мембран, М., 2010.
.Маленков А.Г. и Чуич Г.А. Межклеточные контакты и реакции ткани, М., 2011.
.Сим Э. Биохимия мембран, пер. с англ., М., 2005.
.Финеан Дж., Колмэн Р. и Митчелл Р. Мембраны и их функции в клетке, пер. с англ., М., 2007.
8.Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991-96 гг.
9.Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994. т. 3.
.Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982-1984 гг.
diplomba.ru
1)Биологические мембраны, их структурные компоненты. Функции биологических мембран.
Химический состав и строение биологических мембран.Состав М.б. зависит от их типа и функций, однако основными составляющими являютсяЛипидыиБелки,а такжеУглеводы (небольшая, но чрезвычайно важная часть) и вода (более 20% общего веса).
Липиды.В составе М.б. обнаружены липиды трех классов: фосфолипиды, гликолипиды и стероиды. В мембранах животных клеток более 50% всех липидов составляют фосфолипиды — глицерофосфолипиды (фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит) и сфингофосфолипиды (производные церамида, сфингомиелин). Гликолипиды представлены цереброзидами, сульфатидами и ганглиозидами, а стероиды — в основном холестерином (около 30%). Основную структурную роль в биологических мембранах играют фосфолипиды. Они обладают выраженной способностью формировать бислои при смешивании с водой, что обусловлено химической структурой фосфолипидов, молекулы которых состоят из гидрофильной части — «головки» (остаток фосфорной кислоты и присоединенная к нему полярная группа, например холин) и гидрофобной части — «хвоста» (как правило, две жирно-кислотные цепи). В водной среде фосфолипиды бислоя расположены таким образом, что жирно-кислотные остатки обращены внутрь бислоя и, следовательно, изолированы от окружающей среды, а гидрофильные «головки» —наоборот, наружу.
Холестерин в составе М.б. играет роль модификатора бислоя, придавая ему определенную жесткость за счет увеличения плотности «упаковки» молекул фосфолипидов.
Гликолипиды несут разнообразные функции: отвечают за рецепцию некоторых биологически активных веществ, участвуют в дифференцировке ткани, определяют видовую специфичность.
Белки биологических мембран исключительно разнообразны. Молекулярная масса их в большинстве своем составляет 25 000 — 230 000.
Белки мембран интегральные, периферические.
Рецепторные белки, специфически связывая низкомолекулярные вещества (многие гормоны, медиаторы), обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка принимает различные сигналы, поступающие из внешней среды.
Структурные белки — белки цитоскелета, прилегающие к цитоплазматической стороне клеточной мембраны. В комплексе с микротрубочками и микрофиламентами цитоскелета они обеспечивают противодействие клетки изменению ее объема и создают эластичность.
Углеводыпредставляют собой олиго- или полисахаридные структуры, в состав которых входят глюкоза, галактоза, нейраминовая кислота, фукоза и манноза. Их функции связаны с контролем за межклеточным взаимодействием, поддержанием иммунного статуса клетки, обеспечением стабильности белковых молекул в М.б
Функции биологических мембран.
Барьерная функция. Для клеток и субклеточных частиц М.б. служат механическим барьером, отделяющим их от внешнего пространства.
Перенос веществчерез М.б. сопряжен с такими важнейшими биологическими явлениями, как внутриклеточный гомеостаз ионов, биоэлектрические потенциалы, возбуждение и проведение нервного импульса, запасание и трансформация энергии и т.п. (см.Биоэнергетика). Различают пассивный и активный транспорт (перенос) нейтральных молекул, воды и ионов через М.б.
Способность генерировать биоэлектрические потенциалы и проводить возбуждение. Возникновение биоэлектрических потенциалов связано с особенностями строения биологических мембран и с деятельностью их транспортных систем, создающих неравномерное распределение ионов по обе стороны мембраны.
Процессы трансформации и запасания энергиипротекают в специализированных М.б. и занимают центральное место в энергетическом обеспечении живых систем. Два основных процесса энергообразования — фотосинтез и тканевое дыхание — локализованы в мембранах внутриклеточных органелл высших организмов, а у бактерий — в клеточной (плазматической) мембране.
Метаболические функциимембран определяются двумя факторами: во-первых, связью большого числа ферментов и ферментативных систем с мембранами, во-вторых, способностью мембран физически разделять клетку на отдельные отсеки, отграничивая друг от друга метаболические процессы, протекающие в них.
Клеточная рецепция и межклеточные взаимодействия. Под этой формулировкой объединен весьма обширный и разнообразный набор важных функций клеточных мембран, определяющих взаимодействие клетки с окружающей средой и формирование многоклеточного организма как единого целого. Молекулярно-мембранные аспекты клеточной рецепции и межклеточных взаимодействий касаются прежде всего иммунных реакций, гормонального контроля роста и метаболизма, закономерностей эмбрионального развития
studfiles.net
Основные функции биологических мембран — fiziku5.ru
3) фазового состояния мембранных липидов.
Отклонение биофизических характеристик липидного бислоя от нормы связано с разного рода патологиями. Важную роль в физиологии клетки играют фазовые переходы в биологических мембранах.
По структуре мембрана относится к жидким кристаллам, но ее неверно было бы представлять как застывшую структуру. На поверхностях бислоев происходит интенсивное тепловое движение. Его называют латеральной диффузией. Оно может быть представлено как последовательность перескоков молекул из одного положения в другое с частотой примерно 107—108 в секунду (примерно за это время молекула огибает всю поверхность мембраны). Гораздо реже происходят перескоки молекул с одной на другую поверхность мембраны. Для фосфолипидов этот процесс называется «флип-флоп» и происходит 1 раз в несколько часов.
Плотность липидного бислоя составляет 800 кг/м3, что меньше, чем у воды.
Размеры. По данным электронной микроскопии, толщина мембраны варьирует от 4 до 13 нм, причем различным клеточным мембранам присуща разная толщина.
Прочность. Предел прочности на разрыв для мембраны низок. В условиях организма средние деформации составляют около 0,01 %. Чтобы довести мембрану до разрыва, достаточно внутреннего давления 100 Па. Живая клетка может осуществлять осморегуляцию только за счет изменения своей формы, но не за счет растяжения мембраны.
Деформируемость. Клеточная мембрана легко подвергается деформации сдвига. Например, в потоке эритроцитов с градиентом скорости происходит вращение мембраны вокруг содержимого клетки. Это явление получило название «феномена гусеницы танка». Мембрана обладает высокой гибкостью. При оценке механических свойств мембраны эффективный модуль упругости принимается равным 0,45 Па.
Вязкость. Липидный слой мембраны имеет вязкость η=30-100 мПа∙с (что соответствует вязкости растительного масла). Поверхностное натяжение равно 0,03—3 мН∙ м-1, что на 2—3 порядка ниже, чем у воды (73 мН∙м-1). Коэффициент проницаемости мембранного вещества для воды равен (25-33)∙10-4 см/с.
Мембрана — конденсатор. Двойной фосфолипидный слой уподобляет мембрану плоскому конденсатору, обкладки которого образованы электролитами внеклеточного и внутриклеточного (цитоплазмы) растворами с погруженными в них поверхностными белками и головками липидных молекул. Обкладки разделены диэлектрическим слоем, образованным неполярной частью липидных молекул — двойным слоем их хвостов. Электроемкость 1см2 мембраны составляет 0,5—1,3 мкФ. Напряженность электрического поля в мембране составляет приблизительно 20∙106 В/м. Диэлектрическая проницаемость мембраны составляет: для фосфолипидной области ε = 2,0—2,2; для гидрофильной области ε = 10—20. Электросопротивление 1 см2 поверхности мембраны составляет 102—105 Ом (что в десятки миллионов раз больше сопротивления внеклеточной жидкости или цитоплазмы). Электроизоляционные свойства мембраны значительно превосходят свойства технических изоляторов.
Жидкокристаллическое состояние. Молекулы в мембране размещены не беспорядочно, в их расположении наблюдается дальний порядок. Фосфолипидные молекулы находятся в двойном слое, а их гидрофобные хвосты приблизительно параллельны друг другу. Есть порядок и в ориентации полярных гидрофильных головок. Физическое состояние, при котором есть дальний порядок во взаимной ориентации и расположении молекул, но агрегатное состояние жидкое, называется жидкокристаллическим состоянием.
Жидкокристаллические структуры очень чувствительны к изменению температуры. В мембранных фосфолипидах при понижении температуры происходит переход из жидкокристаллического в гель-состояние. При этом изменяется взаимное положение гидрофобных хвостов (рис. 5) и увеличивается толщина двойного слоя.
Рис.8. Образование плоской бислойной мембраны
Плоские липидные мембраны, наряду с липосомами, широко используются в качестве моделей для изучения электрических свойств мембраны, их проницаемости и других научных исследований. С помощью модельных мембран изучают ряд функций биологических мембран, а том числе, барьерную (например, селективность проницаемости — хорошую проницаемость для воды и плохую для ионов). Можно моделировать биологический транспорт, вводя в модельную мембрану молекулы-переносчики или молекулы каналоформеры.
4. Основные функции биологических мембран
Единство автономности от окружающей среды и одновременно тесной связи с окружающей средой — необходимое условие функционирования живых организмов на всех уровнях их организации, поэтому важнейшее условие существования клетки и, следовательно, жизни — нормальное функционирование биологических мембран.
В табл. 4 перечислены функции цитоплазматических и некоторых внутриклеточных мембран.
Таблица 4. Некоторые функции биологических мембран
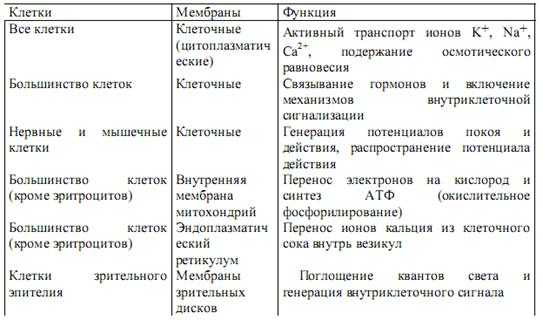
Во всех живых клетках биологические мембрану выполняют функцию барьера, отделяющего клетку от окружающей среды, и разделяющего внутренний объем клетки на сравнительно изолированные «отсеки» (compartments). Сами по себе перегородки, разделяющие клетки на отсеки, построены из двойного слоя липидных молекул и практически непроницаемы для ионов и полярных молекул, растворимых в воде. Но в этот липидный бислой встроены многочисленные белковые молекулы и молекулярные комплексы, одни из которых обладают свойствами селективных (т. е. избирательных) каналов для ионов и молекул, а другие — насосов, способных активно перекачивать ионы через мембрану. Барьерные свойства мембран и работа мембранных насосов создают неравновесное распределение ионов между клеткой и внеклеточной средой, что лежит в основе процессов внутриклеточной регуляции и передачи сигналов в форме электрического импульса между клетками.
fiziku5.ru
Строение и механизмы действия биологической мембраны
Одной из основных особенностей всех эукариотических клеток является изобилие и сложность строения внутренних мембран. Мембраны отграничивают цитоплазму от окружающей среды, а также формируют оболочки ядер, митохондрий и пластид. Они образуют лабиринт эндр-плазматического ретикулума и уплощенных пузырьков в виде стопки, составляющих комплекс Гольджи. Мембраны образуют лизосомы, крупные и мелкие вакуоли растительных и грибных клеток, пульсирующие вакуоли простейших. Все эти структуры представляют собой компартменты (отсеки), предназначенные для тех или иных специализированных процессов и циклов. Следовательно, без мембран существование клетки невозможно.
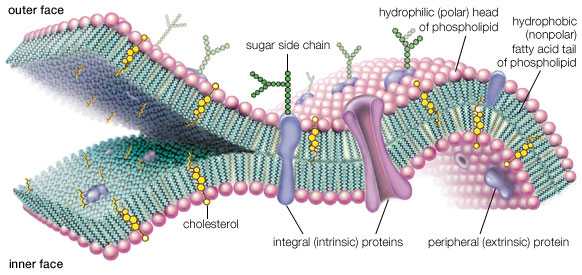
Строение биологической мембраны
Плазматическая мембрана, или плазмалемма, — наиболее постоянная, основная, универсальная для всех клеток мембрана. Она представляет собой тончайшую (около 10 нм) пленку, покрывающую всю клетку. Плазмалемма состоит из молекул белков и фосфолипидов.
Строение биологической мембраны, как основного строительного компонента клетки
Любая живая клетка, в том числе и растительная, имеет сложное строение, состоит из внешней оболочки (клеточной стенки), протоплазмы или цитоплазмы и различных органоидов или органелл, находящихся внутри клетки. Все структурные элементы клетки состоят из биологических мембран, за исключением рибосом. Рибосомы по своему строению не относятся к мембранным органоидам, однако, поскольку они расположены непосредственно на эндоплазматической сети, то именно ЭПС выполняет все функции мембраны по отношению к ним.
Эволюция представлений о строении мембран происходит более 100 лет ( демонстрировать таблицу). Сложность строения этого важнейшего элемента живой клетки позволяет предположить, что в отдельных случаях структура мембран в том или другом конкретном случае может варьировать в пределах предложенных моделей. Более того, окончательной ясности в вопросе о структуре мембраны пока не достигнуто.
Состав мембран зависит от их типа и функции, однако во всех случаях их основными составляющими являются липиды и белки, соотношение между которыми колеблется в пределах от 0.4 до 2.5 Толщина мембран обычно составляет 4-10 нм.
Белковые компоненты мембран состоят из молекул с молекулярной массой от 5000 до 250000. Липидная часть состоит в основном из фосфолипидов, сфинголипидов и стероидов. Электронно-микроскопические исследования показывают наличие трех слоев, из которых два внешних поглощают электроны, а третий, внутренний, их пропускает.
В структуре мембраны согласно жидкостно-мозаичной модели имеется двойной липидный слой, покрытый периферическими белками, кроме того в него погружаются полностью или частично интегральные белки. Эти белки имеют двойственную природу, причем спиральные участки, пронизывающие липидный слой, состоят из алифатических (липофильных) аминокислот, в то время как их наружные концы гидрофильны и могут быть связаны с остатками сахаров.
Основные функции биологических мембран заключаются в: отделении клеток от межклеточной жидкости, создании внутренней архитектуры клетки, в поддержании градиента концентраций и электрохимического градиента, в осуществлении переноса питательных веществ и продуктов жизнедеятельности.
Механизм действия биологической мембраны.
Одной из важнейших функций мембраны является пропускание веществ - обеспечение обмена веществ между клеткой и окружающей средой. Перенос веществ через биологическую мембрану у многоклеточных организмов сводится к трем основным механизмам: пассивная диффузия, облегченная диффузия, активный транспорт.
При пассивном передвижении ионов этот процесс определяется уравнением:
М = м + nFe,
где м — химический потенциал клетки, е — электрический потенциал клетки, М — электрохимический потенциал, n — валентность иона, F — константа Фарадея.
Кроме проникновения в клетку сквозь мембрану, вещества могут поступать в клетку путем пиноцитоза, поверхностной адсорбции. При этом адсорбция может быть:
- физическая или неполярная (когда действуют силы Ван дер Ваальса),
- полярная (адсорбция электронов или ионов),
- хемосорбция (химическое взаимодействие).
Для объяснения механизма облегченной и активной диффузии Беннет-Кларком, Курсановым и Стейном предложена теория клеточных переносчиков, суть которой состоит в том, что в цитоплазме ионы связываются специальными переносчиками и переносятся во внутреннюю часть протопласта (использовать схему).
Связь между клетками, обмен веществ с окружающей средой, осуществляется через клеточную стенку, которая в силу особенностей своего строения играет особую роль в этих процессах. Она не препятствует диффузии, играет роль адсорбента некоторых веществ, имеет в своей структуре особые поры, то есть при необходимости облегчает перенос веществ.
Связь клеток в растительных тканях осуществляется либо по симпластическому, либо по апопластическому типу. При этом в апопласте перенос веществ осуществляется пассивно по межклетникам и активно между межклетниками и клетками. Типичная апопластическая ткань — ксилема.
В симпласте клетки связаны между собой плазмодесмами, которые обеспечивают пассивный перенос веществ между клетками, а активный перенос отмечается между вакуолью и цитоплазмой. Типичная симпластическая ткань - флоэма.
biofile.ru
Мембраны биологические основные функции — Справочник химика 21
Все биологические мембраны, включая плазматическую мембран и внутренние мембраны эукариотических клеток, имеют общие структурные особенности они представляют собой ансамбли липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий. Благодаря этим взаимодействиям поддерживается структурная целостность мембран Однако сами по себе клеточные мембраны являются подвижными, текучими структурами и большинство входящих в их состав молекул способны перемещаться в плоскости мембраны. Как показано на рис. 6-1, липидные молекулы образуют непрерывный двойной слой толщиной около 5 нм. Липидный бислой — это основная структура мембраны, которая и создает относительно непроницаемый барьер для большинства водорастворимых молекул. Белковые молекулы как бы растворены в липидном бислое. С их помощью выполняются разнообразные функции мембраны. Одни мембранные белки обеспечивают транспорт молекул внутрь клетки или из нее, другие являются ферментами и катализируют ассоциированные с мембраной реакции. Еще один класс белков осуществляет структурную связь плазматической мембраны с цитоскелетом, с одной стороны, и(или) с внеклеточным матриксом либо с соседней клеткой — с другой. Отдельную группу составляют белки, выполняющие роль рецепторов для получения и преобразования химических сигналов из окружающей среды. Как и следовало ожидать, мембраны асимметричны оба их слоя различаются по липидному и белковому составу, что отражает, по-видимому, функциональные различия их поверхностей. [c.349]Одно из самых значительных достижений рентгеноструктурного анализа белков последних лет, которое не может не повлиять на дальнейшее развитие биологии и становление ее новой области -молекулярной биологии клетки, состоит в начавшейся расшифровке трехмерных структур первых мембранных белков. Перед обсуждением полученных здесь результатов целесообразно кратко сообщить о том, что было известно об этих белках до исследования их с помощью рентгеновской дифракции. Если основные структурные особенности биологических мембран определяются молекулами липидного бислоя, то специфические функции мембран выполняются главным образом белками. Они ответственны за процессы превращения энергии, выступают в качестве рецепторов и ферментов, образуют каналы активного и пассивного транспорта молекул и ионов различных веществ через мембраны, охраняют организм от проникновения чужеродных антигенов и стимулируют иммунный ответ клеточного типа. В обычной плазматической мембране белок составляет около 50% ее массы. Однако в некоторых мембранах, например во внутренних мембранах митохондрий и хлоропластов, его содержание поднимается до 75%, а в других, например миелиновой мембране, снижается до 25%. Многие мембранные белки пронизывают липидный бислой насквозь и контактируют с водной средой по обеим сторонам мембраны. Молекулы этих белков, называемых трансмембранными, как и окружающие их молекулы липидов, обладают амфипатическими свойствами, поскольку содержат гидрофобные участки, взаимодействующие внутри бислоя с гидрофобными хвостами липидов, и гидрофильные участки, обращенные к воде с обеих сторон мембраны. Другая группа мембранных белков соприкасается с водой только с одной стороны бислоя [234, 235]. Одни из них погружены только во внешний или во внутренний слой мембраны, другие ассоциированы за счет невалентных взаимодействий с трансмембранными белками, третьи прикреплены к мембране с помощью ковалентно связанных с ними цепей жирных кислот, внедренных в липидный слой. [c.56]
Чувствительность биологических процессов к pH обусловлена целым рядом причин.. Ионы водорода могут выступать в качестве катализатора ряда процессов, быть реагентом или продуктом реакции. Кроме того, при изменении pH может измениться проницаемость клеточной мембраны, а следовательно, и распределение веществ или ионов по обе ее стороны. Подобно другим биологическим структурам, мембраны содержат способные к ионизации группы, и в зависимости от степени их ионизации меняется конформация, а значит и биологическая активность молекул, в которые эти группы входят. Это прежде всего касается белков, а следовательно, ферментов. В некоторых белках небольшое изменение pH окружающей среды вызывает проявление биологической активности. На примере гемоглобина, основной функцией которого является перенос кислорода от легких к тканям, можно видеть, что при активном тканевом дыхании незначительное понижение pH в тканях в результате образования углекислоты и ионов водорода облегчает высвобождение кислорода. Процесс высвобождения кислорода сопровождается связыванием протонов гемоглобином, что увеличивает буферную емкость системы. [c.19]
Транспорт через клеточные мембраны основного продукта гидролиза углеводов пищи — глюкозы — регулируется инсулином (гормоном, вырабатываемым поджелудочной железой), строение и биологические функции которого рассмотрены в главе 9. [c.398]
Несмотря на то что каждому типу мембран присущи определенные липидные и белковые компоненты, основные структурные и функциональные особенности, обсуждаемые в этой главе, характерны как для внутриклеточных, так и для плазматических мембран. Прежде всего нам хотелось бы рассмотреть структуру и организацию главных компонентов всех биологических мембран — липидов, белков и углеводов. Затем мы обсудим механизмы, используемые клетками для транспорта малых молекул через плазматическую мембрану, а также способы поглощения и выделения клетками макромолекул и крупных частиц. В последующих главах будут проанализированы некоторые дополнительные функции плазматической мембраны роль в клеточной адгезии (гл. 14) и в сигнальных функциях (гл. 12). [c.349]
Липидный бислой определяет основные структурные особенности биологических мембран, тогда как белки ответственны за большинство мембранных функций. Они выступают в качестве специфических рецепторов и ферментов, осуществляют транспорт через мембрану различных веществ и т. д. Большинство мембранных белков пронизывает бислой в виде одиночной а-спирали но есть и такие, которые пересекают бислой несколько раз в виде серии а-спиралей. Следующая группа белков ассоциирует с мембраной, не пересекая бислой, а прикрепляясь к той или другой стороне мембраны. Многие из этих белков связаны нековалентными взаимодействиями с трансмембранными белками, есть и такие, которые [c.376]
В этой главе мы сначала рассмотрим фундаментальные термодинамические законы, в соответствии с которыми происходит распределение малых молекул по разные стороны полупроницаемой мембраны, и обсудим, как с их помощью можно подойти к рещению некоторых интересных проблем. Эти законы лежат в основе поведения всех полупроницаемых перегородок, в том числе и сложных биологических мембран. Затем мы рассмотрим свойства искусственных биологических мембран — фосфолипидных би
www.chem21.info