Клеточные мембраны » mozok.click
Повторите материал предыдущей темы и объясните, какие органические вещества составляют основу биологических мембран. вспомните, какие свойства присущи липидам. Каких липидов в клеточных мембранах больше всего?
Клеточные мембраны
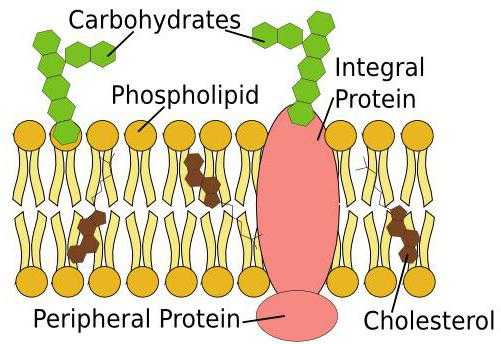
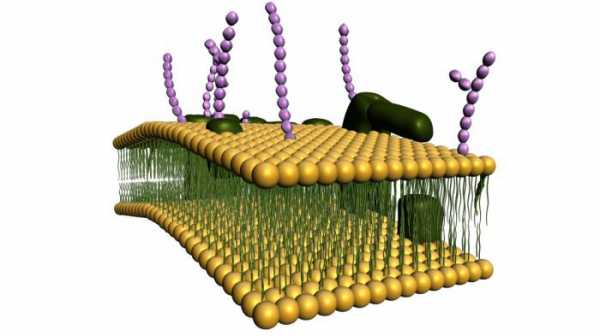
Клеточные мембраны — это структуры, которые окружают клетку и формируют внутри нее ряд органелл. Это так называемые мембранные органеллы — ядро, митохондрии, вакуоли и др. Мембраны образованы двумя слоями липидов, в которых расположены молекулы белков. Белки и липиды мембран нередко присоединяют к себе молекулы углеводов, образуя гликопротеиды и гликолипиды (рис. 10.1). Так как липидные слои, образующие мембрану, жидкообразны и текучи, то погруженные в них белки довольно подвижны. Поэтому модель, которая описывает строение мембраны, называют жидкостно-мозаичной.
Функции мембран
Важнейшими функциями биологических мембран являются барьерная, рецепторная и транспортная. Мембраны являются барьерами с избирательной проницаемостью. Они регулируют обмен веществ между клеткой и окружающей средой, а также между отдельными компонентами внутри клетки. Барьерные функции мембран выполняют липиды. Именно они образуют основу биологической мембраны.
Рецепторную функцию выполняет надмембранный комплекс. Это, главным образом, гликопротеиды, которые образуют структуры, расположенные на поверхности мембраны.
Мембраны играют важную роль в обмене веществ между клеткой и окружающей средой и обеспечивают межклеточные взаимодействия. Именно они передают сигналы из внешней среды клеток во внутреннюю.
Мембраны выполняют также структурную, защитную и ферментативную функции. Они придают клеткам определенную форму, защищают их от повреждений и объединяют отдельные клетки тканей в единое целое. Ферменты, которые располагаются на поверхности мембран, обеспечивают выполнение ферментативной функции. Например, при пищеварении в кишечнике человека.
Транспорт веществ через мембраны
Транспортная функция мембран чрезвычайно важна для жизнедеятельности клетки. Наибольшую роль в ее выполнении играют белки. Они могут формировать в мембране сквозные каналы или транспортировать некоторые вещества в связанном виде, образуя с ними временные соединения.
Перенос веществ через мембрану может происходить либо без затрат энергии (пассивный транспорт), либо с затратами (активный транспорт). Пассивный транспорт осуществляется из зоны с высокой концентрацией вещества в зону с его низкой концентрацией. А активный транспорт, наоборот, — из зоны с низкой концентрацией в зону с высокой концентрацией.
Через мембрану транспортируются как большие, так и малые молекулы веществ. Перемещение малых молекул происходит с помощью простой диффузии, облегченной диффузии, активного транспорта, а больших — путем экто- и эндоцитоза.
Пасивний транспорт
Путем простой диффузии происходит только пассивный транспорт веществ (кислород, углекислый газ). А путем облегченной диффузии может осуществляться как пассивный, так и активный виды транспорта. Облегченная диффузия происходит через специальные
каналы, расположенные внутри больших белковых молекул. Ее могут выполнять также специальные белки-переносчики. Так транспортируются малые органические молекулы (глюкоза, некоторые аминокислоты и др.).
Очень важным для клеток является транспорт молекул воды через мембраны. Его особенностью является то, что необходимо переносить через мембрану только молекулы воды, препятствуя переносу растворенных в ней веществ. Этот процесс осуществляют специальные мембранные белки аквапорины (рис. 10.2).
Активный транспорт
Активный транспорт через клеточную мембрану осуществляется с помощью специальных белковых комплексов, а также путем экзо-или эндоцитоза. Так перемещаются ионы и большие молекулы, для которых мембрана является непроницаемой.
Мембранный транспорт в клетку называется эндоцитоз. Мембранный транспорт из клетки — экзоцитоз (рис. 10.3). Транспорт твердых частиц — это фагоцитоз, а транспорт жидкостей и капель — пиноцитоз.
Примером активного транспорта веществ через мембрану клетки является работа натрий-калиевого насоса. Клетке для нормального функционирования необходимо поддерживать определенное соотношение ионов Na+ и K+ — в цитоплазме и внеклеточной среде.
Перенос ионов Na+ и K+ осуществляется белком, который расположен в клеточной мембране. Этот белок перекачивает ионы Na+ из клетки, а ионы K+ — в клетку. Перенос этих ионов происходит из области их низкой концентрации в область их высокой концентрации (ионов K+ больше в клетке, а ионов Na+ — вне клетки). Для этого процесса используется энергия молекул АТФ.
За один цикл работы насос выкачивает из клетки 3Na+ и закачивает 2K+ (рис. 10.4).

Поэтому на внешней стороне мембраны накапливается избыток положительных ионов. Это создает разность потенциалов между внешней и внутренней поверхностями мембраны. Эту разность используют как источник энергии многие другие белковые комплексы для переноса различных веществ через мембрану.
Клеточные мембраны — это структуры, которые окружают клетку и формируют внутри нее ряд органелл. в их состав входят липиды, белки и углеводы. Липиды образуют двойной слой. в каждом из слоев гидрофильные части липидных молекул ориентированы наружу, а гидрофобные — внутрь. Белковые комплексы в составе мембраны могут пронизывать ее насквозь, размещаться в пределах одного липидного слоя или даже снаружи мембраны. Состав внешнего и внутреннего липидных слоев может различаться. Основными функциями мембран являются барьерная, транспортная и рецепторная.
Проверьте свои знания
1. Что такое мембрана? Какие объекты она окружает? 2. Каково строение клеточной мембраны? 3. Какие вещества входят в состав мембран? 4. Какие функции выполняют клеточные мембраны? 5. Почему некоторые вещества не могут проникать через мембрану путем простой диффузии? 6*. Какие свойства фосфолипидов привели к тому, что они стали основой клеточных мембран? 7*. Почему для разных веществ используются различные механизмы транспорта через мембрану клетки?
Это материал учебника Биология 9 класс Задорожный
Особенности, строение и функции клеточных мембран
В 1972 году была выдвинута теория, согласно которой частично проницаемая мембрана окружает клетку и выполняет ряд жизненно важных задач, а строение и функции клеточных мембран являются значимыми вопросами касательно правильного функционирования всех клеток в организме. Клеточная теория получила широкое распространение в 17 веке, вместе с изобретением микроскопа. Стало известно, что растительные и животные ткани состоят из клеток, но из-за низкой разрешающей способности прибора невозможно было увидеть какие-то барьеры вокруг животной клетки. В 20-м веке химическая природа мембраны исследовалась более детально, было выяснено, что ее основу составляют липиды.

Строение и функции клеточных мембран
Клеточная мембрана окружает цитоплазму живых клеток, физически отделяя внутриклеточные компоненты от внешней среды. Грибы, бактерии и растения также имеют клеточные стенки, которые обеспечивают защиту и препятствуют прохождению крупных молекул. Клеточные мембраны также играют роль в становлении цитоскелета и прикреплении к внеклеточному матриксу других жизненно важных частиц. Это нужно для того, чтобы удерживать их вместе, формируя ткани и органы организма. Особенности строения клеточной мембраны включают проницаемость. Основной функцией является защита. Мембрана состоит из фосфолипидного слоя со встроенными белками. Эта часть участвует в таких процессах, как клеточная адгезия, ионная проводимость и сигнальные системы и служит в качестве поверхности крепления для нескольких внеклеточных структур, в том числе стенки, гликокаликса и внутреннего цитоскелета. Мембрана также сохраняет потенциал клетки, работая как селективный фильтр. Она является селективно проницаемой для ионов и органических молекул и управляет перемещением частиц.
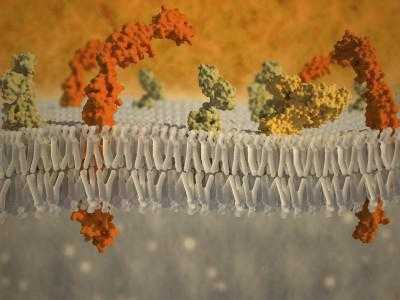
Биологические механизмы с участием клеточной мембраны
1. Пассивная диффузия: некоторые вещества (малые молекулы, ионы), такие как двуокись углерода (СО2) и кислорода (О2), могут проникать через плазматическую мембрану путем диффузии. Оболочка действует как барьер для определенных молекул и ионов, они могут концентрироваться по обе стороны.
2. Трансмембранный белок каналов и транспортеров: питательные вещества, такие как глюкоза или аминокислоты, должны попасть в клетку, а некоторые продукты обмена веществ должны ее покинуть.
3. Эндоцитоз — это процесс, при котором поглощаются молекулы. В плазматической мембране создается небольшая деформация (инвагинация), в которой вещество, подлежащее транспортировке, заглатывается. Это требует энергии и, таким образом, является формой активного транспорта.
4. Экзоцитоз: происходит в различных клетках для удаления непереваренных остатков веществ, принесенных эндоцитозом, чтобы секретировать вещества, такие как гормоны и ферменты, и транспортировать вещество полностью через клеточный барьер.

Молекулярная структура
Клеточная мембрана — это биологическая оболочка, состоящая преимущественно из фосфолипидов и отделяющая содержание всей клетки от внешней среды. Процесс образования происходит самопроизвольно при нормальных условиях. Чтобы понять этот процесс и правильно описать строение и функции клеточных мембран, а также свойства, необходимо оценить характер фосфолипидных структур, для которых является свойственной структурная поляризация. Когда фосфолипиды в водной среде цитоплазмы достигают критической концентрации, они объединяются в мицеллы, которые являются более стабильными в водной среде.

Мембранные свойства
- Стабильность. Это значит, что после образования распад мембраны является маловероятным.
- Прочность. Липидная оболочка достаточно надежная, чтобы предотвратить прохождение полярного вещества, через образованную границу не могут пройти как растворенные вещества (ионы, глюкоза, аминокислоты), так и гораздо более крупные молекулы (белки).
- Динамичный характер. Это, пожалуй, наиболее важное свойство, если рассматривать строение клетки. Клеточная мембрана может подвергаться различным деформациям, может складываться и сгибаться и при этом не разрушиться. При особых обстоятельствах, например, при слиянии везикул или бутонизации, она может быть нарушена, но только на время. При комнатной температуре ее липидные составляющие находятся в постоянном, хаотическом движении, образуя стабильную текучую границу.
Жидкая мозаичная модель
Говоря про строение и функции клеточных мембран, важно отметить, что в современном представлении мембрана как жидкая мозаичная модель, была рассмотрена в 1972 году учеными Сингером и Николсоном. Их теория отражает три основные особенности структуры мембраны. Интегральные мембранные белки способствуют мозаичным шаблоном для мембраны, и они способны на боковое движение в плоскости из-за изменчивой природы липидной организации. Трансмембранные белки являются также потенциально мобильными. Важной особенностью структуры мембраны является ее асимметрия. Что представляет собой строение клетки? Клеточная мембрана, ядро, белки и так далее. Клетка является основной единицей жизни, и все организмы состоят из одной или многих клеток, каждая их которых имеет естественный барьер, отделяющий ее от окружающей среды. Эта внешняя граница ячейки также называется плазматической мембраной. Она состоит из четырех различных типов молекул: фосфолипиды, холестерин, белки и углеводы. Жидкая мозаичная модель описывает структуру клеточной мембраны следующим образом: гибкая и эластичная, по консистенции напоминает растительное масло, так что все отдельные молекулы просто плавают в жидкой среде, и они все способные двигаться вбок в пределах этой оболочки. Мозаика представляет собой что-то, что содержит много разных деталей. В плазматической мембране она представлена фосфолипидами, молекулами холестерина, белками и углеводами.
Фосфолипиды
Фосфолипиды составляют основную структуру клеточной мембраны. Эти молекулы имеют два различных конца: голову и хвост. Головной конец содержит фосфатную группу и является гидрофильным. Это значит, что он притягивается к молекулам воды. Хвост состоит из водорода и атомов углерода, называемых цепочками жирных кислот. Эти цепи гидрофобны, они не любят смешиваться с молекулами воды. Этот процесс напоминает то, что происходит, когда вы льете растительное масло в воду, то есть оно в ней не растворяется. Особенности строения клеточной мембраны связаны с так называемым липидным бислоем, который состоит из фосфолипидов. Гидрофильные фосфатные головы всегда располагаются там, где есть вода в виде внутриклеточной и внеклеточной жидкости. Гидрофобные хвосты фосфолипидов в мембране организованы таким образом, что держат их подальше от воды.

Холестерин, белки и углеводы
Услышав слово «холестерин», люди обычно думают, что это плохо. Однако на самом деле холестерин является очень важным компонентом клеточных мембран. Его молекулы состоят из четырех колец водорода и атомов углерода. Они гидрофобны и встречаются среди гидрофобных хвостов в липидном би-слое. Их важность заключается в поддержании консистенции, они укрепляют мембраны, предотвращая пересечение. Молекулы холестерина также держат фосфолипидные хвосты от вступления в контакт и твердевания. Это гарантирует текучесть и гибкость. Мембранные белки выполняют функции ферментов по ускорению химических реакций, выступают в качестве рецепторов для специфических молекул или транспортируют вещества через клеточную мембрану.
Углеводы, или сахариды, встречаются только на внеклеточной стороне мембраны клетки. Вместе они образуют гликокаликс. Он обеспечивает амортизацию и защиту плазматической мембраны. На основе структуры и типа углеводов в гликокаликсе организм может распознавать клетки и определять, должны ли они быть там или нет.
Мембранные белки
Строение клеточной мембраны животной клетки невозможно представить без такого значимого компонента, как белок. Несмотря на это, они могут значительно уступать по размерам другой важной составляющей – липидам. Существует три вида основных мембранных белков.
- Интегральные. Они полностью охватывают би-слой, цитоплазму и внеклеточную среду. Они выполняют транспортную и сигнализирующую функцию.
- Периферические. Белки прикрепляются к мембране при помощи электростатических или водородных связей в их цитоплазматических или внеклеточных поверхностях. Они участвуют в основном как средство крепления для интегральных белков.
- Трансмембранные. Они выполняют ферментативную и сигнальную функции, а также модулируют основную структуру липидного би-слоя мембраны.

Функции биологических мембран
Гидрофобный эффект, который регламентирует поведение углеводородов в воде, контролирует структуры, образованные посредством мембранных липидов и мембранных белков. Многие свойства мембран даруются носителями липидных би-слоев, образующими базовую структуру для всех биологических мембран. Интегральные мембранные белки частично спрятаны в липидном би-слое. Трансмембранные белки имеют специализированную организацию аминокислот в их первичной последовательности.
Периферические мембранные белки очень похожи на растворимые, но они также привязаны к мембранам. Специализированные клеточные мембраны имеют специализированные функции клеток. Как строение и функции клеточных мембран оказывают влияние на организм? От того, как устроены биологические мембраны, зависит обеспечение функциональности всего организма. Из внутриклеточных органелл, внеклеточного и межклеточных взаимодействий мембран создаются структуры, необходимых для организации и выполнения биологических функций. Многие структурные и функциональные особенности являются общими для бактерий, эукариотических клеток и оболочечных вирусов. Все биологические мембраны построены на липидном би-слое, что обуславливает наличие ряда общих характеристик. Мембранные белки обладают множеством специфических функций.
- Контролирующая. Плазматические мембраны клеток определяют границы взаимодействия клетки с окружающей средой.
- Транспортная. Внутриклеточные мембраны клеток разделены на несколько функциональных блоков с различной внутренней композицией, каждая из которых поддерживается необходимой транспортной функцией в сочетании с проницаемостью управления.
- Сигнальная трансдукция. Слияние мембран обеспечивает механизм внутриклеточного везикулярного оповещения и препятствования разного рода вирусам свободно проникать в клетку.

Значение и выводы
Строение наружной клеточной мембраны оказывает влияние на весь организм. Она играет важную роль в защите целостности, позволяя проникновение только выбранных веществ. Это также хорошая база для крепления цитоскелета и клеточной стенки, что помогает в сохранении формы клетки. Липиды составляют около 50% массы мембраны большинства клеток, хотя этот показатель варьируется в зависимости от типа мембраны. Строение наружной клеточной мембраны млекопитающих являются более сложным, там содержатся четыре основных фосфолипида. Важным свойством липидных би-слоев является то, что они ведут себя как двумерные жидкости, в которой отдельные молекулы могут свободно вращаться и перемещаться в боковых направлениях. Такая текучесть — это важное свойство мембран, которое определяется в зависимости от температуры и липидного состава. Благодаря углеводородной кольцевой структуре холестерин играет определенную роль в определении текучести мембран. Избирательная проницаемость биологических мембран для малых молекул позволяет клетке контролировать и поддерживать ее внутреннюю структуру.
Рассматривая строение клетки (клеточная мембрана, ядро и так далее), можно сделать вывод о том, что организм – это саморегулирующая система, которая без посторонней помощи не сможет себе навредить и всегда будет искать пути для восстановления, защиты и правильного функционирования каждой клеточки.
fb.ru
Биология для студентов — 09. Структура, свойства и состав биологических мембран
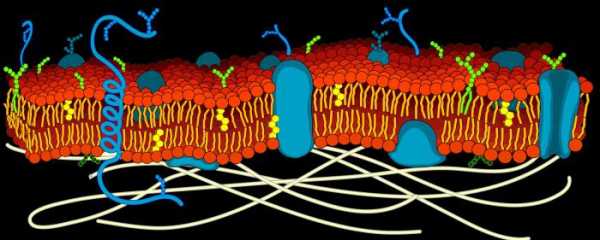
Биологические мембраны — функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму клетки и внутриклеточные органеллы, а также образующие единую внутриклеточную систему канальцев, складок, замкнутых областей. Имеются в клетках всех организмов; отсутствуют только у некоторых вирусов. Мембранные структуры клетки представлены:
- поверхностной (клеточная, или плазматическая) мембраной,
- внутриклеточными мембранами (митохондриальной, ядерной, лизосомной и др.).
Толщина мембраны составляет 7-10 нм, и составляют обычно более половины всей массы клетки (в пересчете на сухой вес). Биологические мембраны состоят в основном из белков, липидов, углеводов и воды. Мембранные липиды — низкомолекулярные вещества. Основную часть липидов — полярные липиды. Они представлены главным образом фосфолипидами — фосфатидилхолином (лецитином) и фосфатидилэтанолами.
Углеводы мембран химически связаны либо с липидами (гликолипиды), либо с белками (гликопротеиды). Гликолипиды и гликопротеиды функционально чрезвычайно важны, поскольку часто определяют иммуноспецифичность клетки, ее способность к взаимодействию с гормонами, медиаторами, токсинами и др. Структурную основу мембран составляет фосфолипидный биомолекулярный слой (бислой), который выполняет функции барьера для ионов и водорастворимых молекул. Он содержит мембранные белки, гликолипиды и гликопротеиды. Белки мембран могут находиться на поверхности липидного бислоя, удерживаемые преимущественно электростатическими силами (периферические белки), либо проникать глубоко в липидный бислой или даже пронизывать его насквозь.
Основные, функции биологических мембран:
- Для клеток и субклеточных частиц мембраны выполняют роль механического осмотического и гидростатического барьера, ограничивающего их от внешнего пространства.
- Одна из центральных функций мембраны. — транспорт веществ через них и регуляция этого процесса.
- Мембрана не статическое образование, а благодаря жидкокристаллической структуре она является двухслойным раствором, в котором липиды способны диффундировать как параллельно поверхности мембраны, так и из одного монослоя в другой.
В структуру мембран обязательно входят белки и их состав варьирует в зависимости от функции той или иной мембраны. В зависимости от прочности связи с мембраной различают:
Периферические,
интегральные белки.
Интегральные белки располагаются между липидами монослоя или пронизывают весь бислой, часто возвышаясь над поверхностью мембраны. Периферические белки связаны с мембранами электростатическими и водородными связями и часто взаимодействуют таким образом с интегральными.
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы «растворены» в липидном бислое. Мембранные липиды — амфифильные (амфипатические) молекулы, т.е. в молекуле есть как гидрофильные группы (полярные «головки»), так и алифатические радикалы (гидрофобные «хвосты»), самопроизвольно формирующие бислой.
Основные принципы структурной организации мембран одинаковы, однако одна из самых характерных особенностей — огромное их разнообразие. Мембраны органелл эукариотических клеток уникальны по своему составу и по характеру выполняемых функций.
Плазматическая мембрана, окружающая каждую клетку:
- определяет её величину,
- обеспечивает транспорт малых и больших молекул из клетки и в клетку,
- поддерживает разницу концентраций ионов по обе стороны мембраны.
Ядерная оболочка состоит из внешней и внутренней ядерных мембран. Ядерная оболочка имеет поры, через которые РНК проникают из ядра в цитоплазму, а регуляторные белки из цитоплазмы в ядро. Мембрана эндоплазматического ретикулума (ЭР) имеет многочисленные складки и изгибы. Она образует непрерывную поверхность, ограничивающую внутреннее пространство, называемое полостью ЭР. Шероховатый ЭР связан с рибосомами, на которых происходит синтез белков плазматической мембраны, ЭР, аппарата Гольджи, лизосом, а также секретируемых белков. Аппарат Гольджи — важная мембранная органелла, отвечающая за модификацию, накопление, сортировку и направление различных веществ в соответствующие внутриклеточные компартменты, а также за пределы клетки.
Митохондриальные мембраны — органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ путём окислительного фосфорилирования. Отличительная особенность внешней митохондриальной мембраны — содержание большого количества белка порина, образующего поры в мембране. Для внутренней мембраны митохондрий характерно высокое содержание белков, около 70%, которые выполняют в основном каталитическую и транспортную функции.
Вода входит в состав мембран и делится на свободную, связанную и захваченную. Связанная и свободная вода различается по подвижности молекул воды и растворяющей способности. Наименьшей подвижностью и растворяющей способностью обладает внутренняя связанная вода. Она присутствует в липидной зоне мембран в виде отдельных молекул. Основную часть связанной воды представляет вода гидратных оболочек. Эта вода окружает полярные группы белков и липидов, имеет min подвижность и практически не обладает свойствами растворителя. Свободная вода в порах и каналах. По ней могут перемещаться свободные ионы. Она является хорошим растворителем, подвижная и обладает всеми свойствами жидкой воды. Захваченная вода обладает изотопным движением, характерным для жидкой воды, является хорошим растворителем. Она встречается в центральной зоне мембран, между ее липидными слоями, но эта вода пространственно делится как с внеклеточной жидкостью, так и с цитоплазмой. У нее нет возможности свободно с ними обмениваться.
Многие мембранные белки и мелкие фрагменты биологических мембран сравнительно легко могут быть включены в состав искусственных везикулярных мембран. Такие комбинированные системы называются протеолипосомами. Эффективность встраивания большинства белков компонентов в искусственные мембранные системы резко зависит от липидного состава мембран, pH, солевого состава, температуры и т. д.
Молекулы липидов, находящиеся в бислое, обладают довольно высокой подвижностью и могут совершать разнообразные движения — поступательные, вращательные и колебательные. В случае липидов большой вклад в подвижность дают внутримолекулярные движения углеводородных цепей. Они происходят путем гош-транс-поворотов смежных звеньев углеводородной цепи вокруг связи С—С. Благодаря высокой конформационной подвижности цепей в них постоянно возникают изгибы и изломы, что приводит к нарушению регулярного расположения липидных молекул в бислое и к появлению в нем дефектов упаковки, называемых «кинки» и «джогги». Наименьшей подвижностью обладает глицериновый остов молекулы, который служит как бы жестким «якорем», ограничивающим движения близлежащих участков углеводородных цепей. По направлению к середине бислоя подвижность цепей возрастает и становится максимальной в области концевых метильных групп. Довольно высокой недвижностью обладает полярная головка липидной молекулы. Помимо движений отдельных участков липидной молекулы относительно друг друга в жидкокристаллическом бислое происходят также движения всей молекулы как единого целого. Они включают:
- аксиальное вращение молекулы вокруг ее длинной оси, перпендикулярной к плоскости бислоя,
- маятниковые и поплавочные колебания молекулы относительно ее равновесного положения в бислое,
- перемещение молекулы вдоль бислоя (латеральная диффузия),
- перескок ее с одной стороны бислоя на другой.
Все эти движения совершаются с разными скоростями. Очень медленно протекает в липидном бислое флип-флоп. Обычно полупериод флип-флопа составляет величины порядка нескольких часов или даже дней.
Многие мембранные белки способны легко диффундировать вдоль мембраны и обладают довольно высокой вращательной подвижностью. Но даже в случае самых подвижных белков измеряемые коэффициентом диффузии примерно на порядок ниже, чем для липидных молекул.
Поляризация биоэлектрическая, возникновение двойного электрического слоя на границе между наружной средой и содержимым живой клетки; при этом наружная поверхность клетки в состоянии покоя заряжена положительно по отношению к её содержимому, имеющему отрицательный заряд. Постоянная биоэлектрическая поляризация обусловлена особенностями строения биологических мембран, а также неравномерным распределением неорганических ионов (в первую очередь К+, Na+, Cl) в содержимом клетки и в окружающей её среде (электрохимические градиенты). Потенциал покоя — непосредственное следствие поляризации.
При исследовании биологических объектов было установлено, что с увеличением частоты тока их электропроводность растет. Зависимость электропроводности от частоты переменного тока получила название дисперсии электропроводности. Диапазон частот проявляющейся дисперсии располагается в интервале 102 – 108 Гц, и характерна эта зависимость для всех тканей Рост электропроводности связан с тем, что при малых частотах проявляются эффекты поляризации, которые по мере увеличения частот переменного тока сказываются меньше. Диэлектрическая проницаемость /ДЭП/ биологических объектов. ДЭП (e) показывает во сколько раз взаимодействие между зарядами в неограниченной однородной среде (e1) меньше, чем в ваккуме (e0)
При пропускании постоянного тока через живые клетки и ткани было установлено, что сила тока не остается постоянной, а сразу же после наложения потенциала начинает непрерывно падать и, наконец, устанавливается на уровне, который во много раз ниже, чем исходный. Это объясняется тем, что при прохождении постоянного тока через биологическую систему в ней возрастает нарастающая до некоторого предела Э.Д.С. противоположного направления.
vseobiology.ru
Характеристика клеточной мембраны (нейролеммы)
Клеточная мембрана – нейролемма – представляет собой тонкую (6 нм) липопротеиновую пластинку, содержание липидов в которой составляет около 40%, белков – около 60%. Изнутри нейролемма выстлана тонким, более плотным слоем гиалоплазмы. На внешней поверхности мембраны имеется небольшое количество углеводов, молекулы которых соединены либо с белками ( гликопротеиды ), либо с липидами ( гликолипиды ) и образуют гликокаликс. Углеводы участвуют в процессах рецепции биологически активных веществ, реакциях иммунитета.
Структурную основу клеточной мембраны — матрикс – составляет бимолекулярный слой фосфолипидов, являющихся барьером для заряженных частиц и молекул водорастворимых веществ. Липиды обеспечивают высокое электрическое сопротивление мембраны.
Молекулы фосфолипидов мембраны состоят из двух частей: одна из них несет заряд и гидрофильна, другая – не заряжена и гидрофобна. Это определяет способность липидов самопроизвольно образовывать двухслойные мембранные структуры под влиянием собственных зарядов. Заряженные гидрофильные участки одних молекул фосфолипидов направлены внутрь, а других – наружу. В толще мембраны молекулы фосфолипидов взаимодействуют незаряженными гидрофобными участками. В липидном слое клеточных мембран находится много холестерина.
Молекулы белков встроены в фосфолипидный матрикс клеточной мембраны. Все белки нейролеммы можно разделить на
а) структурные белки;
б) белки-переносчики;
в) ферменты;
г) каналообразующие белки;
д) ионные насосы;
е) специфические рецепторы.
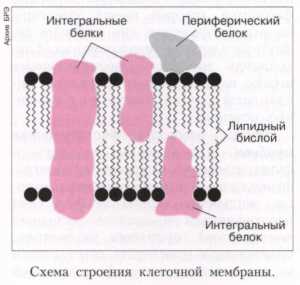
Один и тот же белок может быть рецептором, ферментом и насосом. Многие мембранные белки состоят из двух частей: заряженной и незаряженной. Незаряженные участки белков погружены в липидный слой, не несущий заряда. Заряженные участки белков взаимодействуют с заряженными участками липидов, что является важным фактором, определяющим взаиморасположение структурных элементов клеточной мембраны и ее прочность. Белки, пронизывающие липидный слой, крепко связаны с молекулами фосфолипидов и носят название интегральных белков. Белки, которые располагаются на поверхности мембраны (как снаружи, так и изнутри), называются периферическими белками.
Клеточная мембрана нейрона имеет отрицательный поверхностный заряд, который создается выступающей из мембраны углеводной частью гликолипидов, фосфолипидов и гликопротеидов.
Мембрана обладает текучестью, то есть ее части могут перемещаться из одного участка на другой.
Клеточная мембрана обладает избирательной проницаемостью, то есть одни вещества пропускает, другие не пропускает. В частности нейролемма легко проницаема для жирорастворимых веществ. Анионы органических кислот не проходят через мембрану, но имеют каналы, избирательно пропускающие ионы К, Na, Са, Cl. При действии нервных импульсов проницаемость мембраны нейрона для различных ионов изменяется, это обеспечивает движение ионов согласно концентрационному и электрическому градиентам, что выражается в возникновении возбуждающих и тормозных потенциалов. Проницаемость нейролеммы под действием нервных импульсов может повышаться благодаря наличию в ней ионных управляемых каналов. Каналы образованы белковыми молекулами, встроенными в липидный матрикс. Через эти каналы могут проходить полярные молекулы.
Основные функции клеточной мембраны:
1. Барьерная функция — клеточная мембрана отделяет внутреннюю среду клетки от внешней, защищая, тем самым, клетку от воздействия вредных факторов и поддерживая постоянный состав клетки.
2. Функция восприятия изменений внутренней среды (рецепторная функция) – клеточная мембрана обладает большим набором различных рецепторов, обладающих специфической чувствительностью к различным агентам: гормонам, медиаторам, антигенам, химическим и физическим раздражителям. Рецепторы отвечают за взаимное распознавание клеток, развитие иммунитета. Рецепторами могут служить гликопротеиды и гликопептиды мембраны. Возбужденный рецептор активирует G-белок мембраны, который с помощью фермента-предшественника, расположенного на внутренней стороне мембраны, активирует вторичный посредник, реализующий эффект от раздражителя, подействовавшего на рецептор. Восприятие химических и физических раздражителей у возбудимых клеток осуществляется с помощью трансформации энергии раздражения в нервный импульс.
3. Функция создания электрического заряда клетки — каждая клетка имеет мембранный потенциал - разность концентрации заряженных ионов на внутренней и внешней поверхности мембраны. Благодаря наличию мембранного потенциала ( МП ) возбудимые клетки могут генерировать потенциал действия (возбуждение), и передавать его на другую клетку. Распространение возбуждения обеспечивает быструю связь возбудимых клеток между собой и с эффекторными клетками. МП нейрона варьирует от 60 до 80 мВ, МП клеток нейроглии равен 90 мВ.
4. Функция передачи сигналов от одной клетки к другой – осуществляется с помощью синапсов, образованных в области контакта нейронов друг с другом.
5. Транспортная функция – определяет состав веществ в клетке. Клеточная мембрана осуществляет тонкую регуляцию содержания в цитоплазме ионов и молекул. Благодаря транспорту частиц формируется состав внутриклеточной среды, наиболее благоприятной для оптимального протекания метаболических реакций. Транспорт частиц через клеточную мембрану нейрона обеспечивает:
1) поступление в клетку различных веществ, необходимых для синтеза клеточных структур и выработки энергии;
2) выделение клетками продуктов ее обмена и биологически активных веществ – нейрогормонов, нейромедиаторов;
3) создание электрических зарядов клеток, возникновение и распространение возбуждения.
Первичный транспорт веществ через клеточную мембрануПервичный транспорт – это такой транспорт, при котором энергия расходуется непосредственно на перенос частиц. Он включает, во-первых, перенос отдельных ионов вопреки концентрационному и электрическому градиентам с помощью специальных ионных насосов, во-вторых, эндоцитоз, экзоцитоз и трансцитоз.
► 1. Транспорт веществ с помощью помп. Насосы представляют собой белковые молекулы, обладающие свойствами переносчика и АТФазной активностью. Непосредственным источником энергии является АТФ. Существуют три основных вида насосов: натрий-калиевый, кальциевый и водородный. Предполагают, что существуют еще хлорный насос. Помпы локализуются на клеточных мембранах или мембранах органелл клеток. Все насосы обладают специфичностью, то есть, способны переносить только один или два определенных иона.
· Натрий-калиевый насос (Na+/K+-АТФаза)– это интегральный белок клеточной мембраны, обладающий свойствами фермента, то есть сам переносчик обеспечивает расщепление АТФ и освобождение энергии, которую сам же и использует. Он имеется на мембранах всех клеток и создает характерный признак живого – градиент концентрации Na+ K+ внутри и вне клетки, который обеспечивает формирование МП и вторичный транспорт веществ. Главными активаторами насоса являются гормоны альдостерон и тироксин. Ингибирует насос недостаток энергии – кислородное голодание. Блокаторами насоса являются строфантины.
· Кальциевый насос локализуется в эндоплазматичеком ретикулуме и клеточной мембране, он обеспечивает транспорт Са+ в клетке. Насос строго контролирует содержание Са+ в клетке, поскольку изменение содержания Са+ нарушает ее функционирование. Насос переносит Са+ либо во внеклеточную среду, либо в цистерны ретикулума и митохондрии.
· Протонный насос работает в митохондриях нейрона.
· Хлорный насос работает в мембране, ретикулуме и митохондриях.
Постоянная работа помп необходима для поддержания концентрационных градиентов ионов, связанного с ними электрического заряда клетки, движения воды и незаряженных частиц в клетку и из клетки. Совокупность этих процессов обеспечивает жизнедеятельность нейрона. В результате разной проницаемости клеточной мембраны для отдельных ионов и постоянной работы ионных насосов концентрации различных ионов внутри клетки и снаружи нее неодинакова. Поскольку ионы являются заряженными частицами, то существует электрический заряд нейрона: внутри он заряжен отрицательно, то есть количество отрицательно заряженных частиц преобладает, а снаружи положительно, так как положительно заряженных частиц здесь больше. Ионы калия находятся преимущественно в клетке, а ионы натрия и хлора – во внеклеточной жидкости. Внутри клетки имеются также крупномолекулярные анионы, в основном белкового происхождения. Натрий-калиевый насос способен транспортировать еще глюкозу и аминокислоты.
Механизм работы ионных насосов заключается в следующем: Na+/К+ -насос (молекула интегрального белка) переносит за один цикл 3Na+ из клетки и 2К+ в клетку (антипорт), это осуществляется в результате конформации молекулы белка в форму Е1 или Е2. Молекула имеет активный участок, который связывает либо Na+, либо К+. При конформации Е1 активная часть белковой молекулы обращена внутрь клетки и обладает сродством к Na+, который присоединяется к белку. Вследствие этого активируется его АТФаза, обеспечивающая гидролиз АТФ и высвобождение энергии. Последняя обеспечивает изменение конформации молекулы белка: она превращается в форму Е2, вследствие чего ее активный участок будет обращен наружу клеточной мембраны. Теперь белок теряет сродство к Na+, который отщепляется от него, а белок-помпа приобретает сродство к К+ и соединяется с ним. Это снова ведет к изменению конформации белка: форма Е2 переходит в форму Е1. Переносчик теряет сродство к иону К+, который отщепляется, а белок снова меняет свою конформацию. И цикл повторяется. Насос является электронным, поскольку за один цикл выводится из клетки три иона Na+, а в клетку возвращаются два иона К+. Энергия расходуется только на перенос Na+. На обеспечение одного цикла работы Na+/К+ — помпы расходуется одна молекула АТФ.
Подобным же образом работают и другие насосы. Отличие только составляют ионы, которые переносятся помпой.
► 2. Микровезикулярный транспорт. К этому виду транспорта относят эндоцитох, экзоцитоз и трансцитоз. Эндоцитоз – это перенос частиц в клетку. Эндоцитоз – перенос частиц из клетки. Трансцитоз – перенос веществ через клетку, который для нейрона не характерен. С помощью этих процессов переносятся крупномолекулярные белки, полисахариды, нуклеиновые кислоты.
При эндоцитозе клеточная мембрана образует впячивания внутрь клетки, куда захватываются частицы из вне. Эти впячивания, отшнуровываясь внутри клетки, образуют пузырьки. Последние сливаются с лизосомами, образуя вторичные лизосомы, где содержимое подвергается гидролизу.
Экзоцитоз – процесс обратный эндоцитозу. Экзоцитозные пузырьки образуются в аппарате Гольджи. В пузырьки упаковываются белки, образовавшиеся в рибосомах. Пузырьки транспортируются с помощью сократительного аппарата клетки к клеточной мембране, сливаются с ней и их содержимое выделяется во внешнюю среду. Энергия АТФ в обоих случаях расходуется на деятельность сократительного аппарата клетки.
Вторичный транспорт веществ через нейролеммуВторичный транспорт – это переход различных частиц и молекул воды за счет ранее запасенной энергии (потенциальной). Потенциальная энергия создается в виде электрического и концентрационного градиентов, гидростатического давления, что обеспечивает транспорт веществ через клеточную мембрану нейрона и кровеносных сосудов. К вторичному транспорту относятся все виды диффузии – простая, облегченная и осмос.
Диффузия. Согласно законам диффузии, частицы перемещаются из области с высокой концентрацией в область с низкой концентрацией. Частицы с одноименными электрическими зарядами отталкиваются, с разноименными – притягиваются друг к другу. Направление диффузии определяется взаимодействием электрического и концентрационного (химического) градиентов. Если частицы не заряжены, то направление их диффузии определяется только градиентом концентрации. Скорость диффузии зависит от проницаемости мембраны, а также от градиента концентрации незаряженных частиц, и от градиента электрического и химического для заряженных частиц. Направления действия электрического и химического градиентов могут не совпадать. Тогда переход ионов в клетку обеспечивается химическим градиентом вопреки электрическому. Совокупность химического и электрического градиентов называют электрохимическим градиентом. Различают следующие виды диффузии:
1. Простая диффузия осуществляется либо непосредственно через липидный бислой, либо через каналы. При этом заряженные частицы движутся согласно электрохимическому градиенту, а не заряженные – согласно химическому градиенту. А) Через липидный бислой проходят жирорастворимые частицы. Если они находятся в гидратной оболочке по одну сторону мембраны, то могут внедряться в липидную оболочку благодаря тепловому движению, но только после освобождения от оболочки. Частицы неэлектролиты обычно легко освобождаются от гидратной оболочки. С уменьшением молекулярной массы способность перехода частиц через мембрану возрастает. В) По каналам могут проходить различные частицы, причем скорость перемещения очень высока. Каналы заполнены водой, и кроме ионов через них могут проходить малые молекулы неэлектролитов и заряженные молекулы. Скорость простой диффузии определяется электрохимическим градиентом и проницаемостью мембраны для данного вещества. С течением времени скорость простой диффузии изменяется мало. С помощью простой диффузии осуществляется транспорт молекул алкоголя, кислорода, тироксина, углекислого газа, мочевины, лекарственных препаратов. Этот процесс происходит слишком медленно и плохо контролируется.
2. Облегченная диффузия осуществляется согласно химическому градиенту, но она обеспечивает перенос веществ, способных образовывать комплексы с молекулами-переносчиками. Переносчик – белковая молекула мембраны — свободно совершает челночные движения с одной стороны мембраны на другую, либо встраиваются в мембрану, образуя канал. Этот транспорт осуществляется очень быстро. Движущей силой является градиент транспортируемого вещества. С помощью простой диффузии через мембрану могут проходить такие простые вещества как глюкоза, а сахара и аминокислоты проходят только при образовании канала. Облегченная диффузия, в отличие от простой диффузии, не может идти постоянно, так как в данном случае возможно явление насыщения, то есть перенос замедляется или совсем прекращается, если все переносчики заняты.
3. Осмос – частный случай диффузии. Осмос – это движение воды через полупроницаемую мембрану в область с большей концентрацией частиц, то есть с большим осмотическим давлением. Осмотическое давление – это диффузное давление, обеспечивающее движение растворителя через полупроницаемую мембрану. Осмос продолжается до выравнивания осмотического давления по обе стороны полупроницаемой мембраны или выравнивания осмотического давления и гидростатического противодавления. При подавлении метаболизма клетки быстро набухают, так как внутри клетки осмотическое давление сохраняется повышенным. Вода поступает в клетку через водные каналы и временные поры, которые образуются между молекулами липидов и при смещении белковых молекул. Через водные каналы могут проходить малые незаряженные молекулы кислорода, углекислого газа, этанола, мочевины.
Механизмы вторичного транспортаI. Натриевый механизм: энергия затрачивается на создание градиента натрия. Различают два варианта данного механизма транспорта.
Вариант 1. При этом варианте направление движения транспортируемого вещества совпадает с направлением движения натрия согласно его электрохимическому градиенту (симпорт). Глюкоза соединяется с белком-переносчиком, который соединяется с Na+, а Na+, согласно концентрационному и электрическому градиентам диффундирует в клетку и несет с собой глюкозу. В клетке комплекс распадается, Na+ выводится помпой с непосредственной затратой энергии из клетки в интерстиций вопреки электрохимическому градиенту. Транспорт веществ с помощью натрия осуществляется согласно законам диффузии. Транспортируемое вещество при этом может поступать в клетку вопреки собственному химическому градиенту. Движущей силой является электрохимический градиент Na+. Вместе с Na+ в клетку поступает глюкоза, даже если ее концентрация в клетке больше, чем в среде, и если, конечно, электрохимический градиент Na+ превосходит химический градиент глюкозы. С помощью этого варианта обеспечивается реабсорбция медиатора в пресинаптическую терминаль из синаптической щели, а также транспорт глюкозы.
Вариант 2. При втором варианте натриевого механизма перемещение транспортируемых частиц направлено в противоположную сторону движения Na+ (антипорт). С помощью этого обменного механизма регулируется содержание Са2+ в клетке и рН внутри клетки за счет выведения Н-иона в обмен на внеклеточный Na+. Хотя внутриклеточная концентрация Са2+ на несколько порядков ниже внеклеточной, Са2+ выводится из клетки в обмен на поступающий в нее Na+ и противоположно направленные потоки этих ионов сопряжены друг с другом. Этот процесс обеспечивается натриевого химического градиента переносчиком. Переносчик может транспортировать Са2+ и Н+ вопреки их электрическим и химическим градиентам только в том случае, когда сам переносчик имеет собственный градиент: его концентрация на внешней стороне мембраны больше, чем на внутренней.
II. Транспорт веществ из кровеносных сосудов в интерстиций ЦНС осуществляется с помощью диффузии, осмоса и фильтрации, то есть перехода раствора через полупроницаемую мембрану (стенку сосуда) под действием градиента гидростатического давления между жидкостями по обе стороны мембраны. Градиент гидростатического давления создается либо деятельностью сердца, либо гладкой мускулатурой желудочно-кишечного тракта и мышечного пресса.
Ионные каналыИонные каналы образованы белками, они весьма разнообразны по устройству и механизму действия. Классификация ионных каналов осуществляется по нескольким признакам.
· По возможности управления различают управляемые и неуправляемые каналы. Через неуправляемые каналы ионы перемещаются постоянно, но медленно. Через управляемые каналы ионы могут двигаться и быстро и медленно. Управляемые каналы имеют ворота с механизмами управления. Потенциал действия (ПД) в нейроне возникает в основном вследствие активации быстрых Na+ и К+ -каналов. Через неуправляемые каналы и быстрые управляемые каналы ионы перемещаются согласно электрохимичекому градиенту.
· В зависимости от стимула, активирующего или инактивирующего управляемые ионные каналы, в нейронах ЦНС различают потенциалчувствительные и хемочувствительные каналы. Хемочувствительные каналы открываются в результате конформационных изменений рецепторного комплекса. Ворота потенциалзависимых каналов открываются и закрываются при изменении величины мембранного потенциала.
· В зависимости от селективности различают ионоселективные каналы, пропускающие только один ион, и каналы, не обладающие селективностью. В нейронах имеются Na+, K+, Ca2+ и Cl— -селективные каналы. Есть каналы, пропускающие несколько ионов. Такие каналы называются неселективными. Наиболее высока степень селективности у потенциалчувствительных каналов. Для одного и того же иона может существовать несколько каналов.
Наиболее важными для формирования биопотенциалов являются следующие каналы:
I. Каналы для ионов калия.
· Калиевые неуправляемые каналы покоя – через них постоянно К+ выходит из клетки, что является главным фактором в формировании мембранного потенциала (МП) или потенциала покоя (ПП).
· Потенциалчувствительные управляемые К+ -каналы – сравнительно медленно активируются при возбуждении клетки в фазу деполяризации с последующим увеличением активации, что обеспечивает быстрый выход К+ из клетки и ее реполяризацию (генерация потенциала действия – ПД).
II. Каналы для ионов натрия.
· Медленные неуправляемые каналы – каналы утечки, через которые Na+ постоянно диффундирует в клетку и переносит с собой другие молекулы, например, глюкозу, аминокислоты, молекулы-переносчики. Эти каналы участвуют в формировании МП и обеспечивают вторичный транспорт веществ.
· Быстрые потенциалчувствительные Na-каналы быстро активируются при уменьшении МП, что обеспечивает вход Na+ в клетку при возбуждении (восходящая часть потенциала действия — ПД). Затем эти каналы быстро инактивируются.
Устройство ионных каналов и их функционирование
Каналы имеют устье и селективный фильтр, а управляемые каналы – и воротный механизм. Каналы заполнены жидкостью, их размеры 0,3-0,8 нм. Селективность ионных каналов определяется их размером и наличием в канале заряженных частиц. Заряд частиц противоположен заряду иона, который они притягивают, что обеспечивает проход иона через канал. Через ионные каналы могут проходить и незаряженные частицы. Ионы, проходя через канал, должны избавиться от гидратной оболочки, иначе ее размеры будут больше размеров канала. Слишком маленький ион, проходя через селективный фильтр, не может отдать гидратную оболочку, поэтому он не способен пройти через канал.
Для управляемых каналов существуют свои особенности функционирования: во-первых, они отличаются по степени селективности – наиболее высокой степенью селективности обладают потенциалчувствительные каналы; во-вторых, у них может наблюдаться взаимное влияние друг на друга. Так, деполяризация клеточной мембраны за счет активации хемочувствительных каналов для ионов натрия обеспечивает возбуждение нейрона. В то время как активация потенциалчувствительных каналов не влияет на функцию хемочувствительных каналов нейрона.
Ионные каналы блокируются специфическими веществами и фармакологическими препаратами. Новокаин, например, блокирует Na+-каналы, прекращая проведение возбуждения по нервным волокнам.
biofile.ru
Структура и свойства клеточных мембран
Тема: «СТРУКТУРА И СВОЙСТВА КЛЕТОЧНЫХ МЕМБРАН.
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ»
1. Научно-методическое обоснование темы:
Биологические мембраны являются важной частью клетки. Они ограничивают клетку от окружающей среды, защищают ее от вредных внешних воздействий, управляют обменом веществ между клеткой и ее окружением, способствуют генерации электрических потенциалов. Нарушение функций клеточной и внутриклеточной мембраны лежит в основе необратимого повреждения клеток и, как следствие, развития тяжелых заболеваний сердечно-сосудистой, нервной, эндокринной систем. Поэтому медикам необходимо знать физические свойства биологических мембран и основные физические процессы, которые в них происходят.
Живые системы на всех уровнях организации — открытые системы, поэтому транспорт веществ через биологические мембраны — необходимое условие жизни. С переносом веществ через мембраны связаны процессы метаболизма клетки, биоэнергетические процессы, образование биопотенциалов, генерация нервного импульса и др. Нарушение транспорта веществ через биомембраны приводит к различным патологиям. Лечение часто связано с проникновением лекарств через клеточные мембраны. Эффективность лекарственного препарата в значительной степени зависит от проницаемости для него мембраны.
2. Теория:
1.Структура и строение мембран
В конце XX столетия становилось все более очевидным, что большинство клеточных функций осуществляется при непосредственном участии мембран. Растительные и животные клетки разделены на отсеки, многие цитоплазматические органеллы имеют мембранную природу. Кроме органелл, свойственных большинству клеток, имеются и специализированные мембранные системы, например цитоплазматический ретикулум мышечных клеток, миелиновая оболочка периферических нервных волокон и т. д. Поэтому изучение физических основ функционирования биомембран стало очень важным.
Мембраны представляют собой плоские или изогнутые слои толщиной до 10 нм, образованные молекулами белков, жиров (липидов) и углеводов. Мембраны – это клеточные структуры, повсеместно встречающиеся в живых клетках и регулирующие обмен различными веществами между клеткой и внешней средой (клеточные или плазматические мембраны), либо между различными частями клетки (внутриклеточные мембраны).
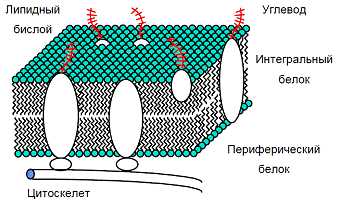
Согласно современным представлениям, все клеточные и внутриклеточные мембраны устроены сходным образом: основу мембраны составляет двойной молекулярный слой липидов (липидный бислой), на котором и в толще которого находятся белки (рис.1). В липидный бислой встроены белковые молекулы, придающие специфические свойства различным участкам мембран, и тем самым, позволяющие последним принимать участие в разнообразных метаболических процессах.
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин (см. табл. 2).
Таблица 2. Состав липидов в мембранах эритроцитов человека
Липидные бислои образуются амфифильными молекулами фосфолипидов и сфингомиелина в водной фазе. Амфифильными эти молекулы называют потому, что они состоят из двух частей, различных по своей растворимости в воде: полярной головы обладающей высоким сродством к воде, т. е. гидрофильной, и хвоста, образуемого неполярными углеводородными цепями жирных кислот; эта часть молекулы обладает низким сродством к воде, т. е. гидрофобна.
Основу (матрицу) мембраны образуют фосфолипидные молекулы, каждая из которых имеет полярную голову и неполярный хвост (рис.1).
Рис.2. Фосфолипидный бислой
Данные рентгеноструктурного анализа показывают, что молекулы фосфолипидов имеют форму сплюснутого с боков цилиндра, а по длине как бы делятся на две неравные части: небольшую «голову», состоящую из полярных групп, и длинный «хвост», образованный углеводородными цепями жирных кислот, входящих в состав фосфолипида (рис.3).
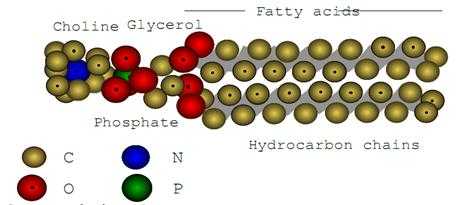
Рис.3. Структура фосфолипида
Именно такое строение молекулы и приводит к тому, что в водных растворах фосфолипидные молекулы самособираются в бислойную мембрану. В мембране «жирные хвосты» упрятаны внутрь, а снаружи в контакте с водным окружением оказываются полярные «головы» этих молекул.
Рис. 4. Самосборка фосфолипидных молекул в липидные везикулы в водном растворе.
В водном растворе происходит самосборка мембран (справа) и замыкание мембран с образованием липидных пузырьков, называемых липосомами (слева).
В настоящее время общепринятой является предложенная в 1972 г. Синджером и Николсоном жидко-мозаичная модель строения биологических мембран. Структурную основу биологической мембраны образует двойной слой фосфолипидов, инкрустированный белками (рис. 5).
Рис.5. Жидко-мозаичная модель мембраны. 1 – поверхностные белки; 2 – полупогруженные белки; 3 –погруженные или интегральные белки; 4 – белки, формирующие ионный канал; 5-ионный канал.
Как упоминалось выше, кроме фосфолипидов и белков, в биологических мембранах содержатся и другие химические соединения. В мембранах животных клеток много холестерина (в сравнимом количестве с фосфолипидами и белками). Есть в мембранах и другие вещества, например гликолипиды, гликопротеиды.
Жидкостно-мозаичная модель строения мембраны, как всякая модель, дает довольно упрощенную картину строения мембраны. В частности выяснилось, что не все липиды в мембране расположены по принципу бислоя. Физические методы исследования показали, что липидная фаза мембран содержит также участки, где липидные молекулы не образуют двойной слой.
Оказалось также, что липидный бислой не является полностью однородным, а содержит домены, названные позднее рафтами (рафт в переводе с английского означает плот или паром). Липидные рафты являются динамическими ансамблями белков и липидов, способными свободно плавать в жидком бислое клеточной мембраны. Как правило, они содержат сфинголипиды и холестерин во внешнем монослое мембраны, связанными с фосфолипидами и холестерином во внутреннем монослое. Обнаружены были рафты благодаря их необычайно высокой устойчивости к действию детергентов, которые легко растворяли липиды, не образующие рафты. Эти структуры
более упорядочены и обладают высокой плотностью благодаря тому, что рафт обычно включает липиды с насыщенными жирнокислотными радикалами. Принципиально важным обстоятельством является то, что в состав рафта включаются многие мембранные белки. В отличие от классической модели Синджера-Николсона белки способны к перемещению в плоскости мембраны по большей части не сами по себе, а в составе рафтов. В физиологии с рафтами связывают функционирование системы внутриклеточной сигнализации. Участие рафтов доказано для многих клеточных патологий, включая болезнь Альцгеймера, болезнь Паркинсона, атеросклероз, бактериальные и вирусные инфекции.
2. Физические свойства мембран. Подвижность фосфолипидных молекул в мембранах
Режим функционирования мембраны сильно зависит от:
1) микровязкости липидного бислоя,
2) подвижности фосфолипидных молекул в мембране,
fiziku5.ru
Плазматическая мембрана: функции, строение :: SYL.ru
Клетка давно определена как структурная единица всего живого. И это действительно так. Ведь миллиарды этих структур, словно кирпичики, образуют растения и животных, бактерий и микроорганизмов, человека. Каждый орган, ткань, система организма — все выстроено из клеток.
Поэтому очень важно знать все тонкости ее внутреннего строения, химического состава и протекающих биохимических реакций. В данной статье рассмотрим, что представляет собой плазматическая мембрана, функции, которые она выполняет, и строение.
Органеллы клетки
Органеллами называются мельчайшие структурные части, находящие внутри клетки и обеспечивающие ее строение и жизнедеятельность. К ним относится множество разных представителей:
- Плазматическая мембрана.
- Ядро и ядрышки с хромосомным материалом.
- Цитоплазма с включениями.
- Лизосомы.
- Митохондрии.
- ЭПС (эндоплазматический ретикулум).
- Комплекс Гольджи.
- Рибосомы.
- Вакуоли и хлоропласты, если клетка растительная.
Каждая из перечисленных структур имеет свое сложное строение, сформирована ВМС (высокомолекулярными веществами), выполняет строго определенные функции и принимает участие в комплексе биохимических реакций, обеспечивающих жизнедеятельность всего организма в целом.

Общее строение мембраны
Строение плазматической мембраны изучалось еще с XVIII века. Именно тогда впервые была обнаружена ее способность выборочно пропускать или задерживать вещества. С развитием микроскопии исследование тонкой структуры и строения мембраны стало более возможным, и поэтому на сегодняшний день о ней известно практически все.
Синонимом ее основному названию является плазмалемма. Состав плазматической мембраны представлен тремя основными видами ВМС:
- белки;
- липиды;
- углеводы.
Соотношение этих соединений и расположение может варьироваться у клеток разных организмов (растительной, животной или бактериальной).
Жидкостно-мозаичная модель строения
Многие ученые пытались высказывать предположения о том, каким образом располагаются липиды и белки в мембране. Однако только в 1972 г. учеными Сингером и Николсоном была предложена актуальная и сегодня модель, отражающая строение плазматической мембраны. Она названа жидкостно-мозаичной, и суть ее состоит в следующем: различные типы липидов располагаются в два слоя, ориентируясь гидрофобными концами молекул внутрь, а гидрофильными наружу. При этом вся структура, подобно мозаике, пронизана неодинаковыми типами белковых молекул, а также небольшим количеством гексоз (углеводов).
Вся предполагаемая система находится в постоянной динамике. Белки способны не просто пронизывать билипидный слой насквозь, но и ориентироваться у одной из его сторон, встраиваясь внутрь. Или вообще свободно «гулять» по мембране, меняя местоположение.
Доказательствами в защиту и оправданность этой теории служат данные микроскопического анализа. На черно-белых фотографиях явно видны слои мембраны, верхний и нижний одинаково темные, а средний более светлый. Также проводился ряд опытов, доказывающих, что слои основаны именно липидами и белками.

Белки плазматической мембраны
Если рассматривать процентное соотношение липидов и белков в мембране растительной клетки, то оно будет примерно одинаковое — 40/40%. В животной плазмалемме до 60% приходится на белки, в бактериальной — до 50%.
Плазматическая мембрана состоит из разных видов белков, и функции каждого из них также специфические.
1. Периферические молекулы. Это такие белки, которые ориентированы на поверхности внутренней или наружной частей бислоя липидов. Основные типы взаимодействий между структурой молекулы и слоем следующие:
- водородные связи;
- ионные взаимодействия или солевые мостики;
- электростатическое притяжение.
Сами периферические белки — растворимые в воде соединения, поэтому их отделить от плазмалеммы без повреждений несложно. Какие вещества относятся к этим структурам? Самое распространенное и многочисленное — фибриллярный белок спектрин. Его в массе всех мембранных белков может быть до 75% у отдельных клеточных плазмалемм.

Зачем они нужны и как зависит от них плазматическая мембрана? Функции следующие:
- формирование цитоскелета клетки;
- поддержание постоянной формы;
- ограничение излишней подвижности интегральных белков;
- координация и осуществление транспорта ионов через плазмолемму;
- могут соединяться с олигосахаридными цепями и участвовать в рецепторной передаче сигналов от мембраны и к ней.
2. Полуинтегральные белки. Такими молекулами называются те, что погружены в липидный бислой полностью или наполовину, на различную глубину. Примерами могут служить бактериородопсин, цитохромоксидаза и другие. Их называют также «заякоренными» белками, то есть будто прикрепленными внутри слоя. С чем они могут контактировать и за счет чего укореняются и удерживаются? Чаще всего благодаря специальным молекулам, которыми могут быть миристиновые или пальмитиновые кислоты, изопрены или стерины. Так, например, в плазмалемме животных встречаются полуинтегральные белки, связанные с холестерином. У растений и бактерий таких пока не обнаружено.
3. Интегральные белки. Одни из самых важных в плазмолемме. Представляют собой структуры, формирующие что-то вроде каналов, пронизывающих оба липидных слоя насквозь. Именно по этим путям осуществляются поступления многих молекул внутрь клетки, таких, которые липиды не пропускают. Поэтому основная роль интегральных структур — формирование ионных каналов для транспорта.
Существует два типа пронизывания липидного слоя:
- монотопное — один раз;
- политопное — в нескольких местах.
К разновидностям интегральных белков можно отнести такие, как гликофорин, протеолипиды, протеогликаны и другие. Все они нерастворимы в воде и тесно встроены в липидный слой, поэтому извлечь их без повреждения структуры плазмалеммы невозможно. По своему строению эти белки глобулярные, гидрофобный конец их расположен внутри липидного слоя, а гидрофильный — над ним, причем может возвышаться над всей структурой. За счет каких взаимодействий интегральные белки удерживаются внутри? В этом им помогают гидрофобные притяжения к радикалам жирных кислот.

Таким образом, существует целый ряд разных белковых молекул, которые включает в себя плазматическая мембрана. Строение и функции этих молекул можно объединить в несколько общих пунктов.
- Структурные периферические белки.
- Каталитические белки-ферменты (полуинтегральные и интегральные).
- Рецепторные (периферические, интегральные).
- Транспортные (интегральные).
Липиды плазмалеммы
Жидкий бислой липидов, которыми представлена плазматическая мембрана, может быть очень подвижным. Дело в том, что разные молекулы могут из верхнего слоя переходить в нижний и наоборот, то есть структура динамична. Такие переходы имеют свое название в науке — «флип-флоп». Образовалось оно от названия фермента, катализирующего процессы перестройки молекул внутри одного монослоя или из верхнего в нижний и обратно, флипазы.
Количество липидов, которое содержит клеточная плазматическая мембрана, примерно такое же, как число белков. Видовое разнообразие широко. Можно выделить такие основные группы:
- фосфолипиды;
- сфингофосполипиды;
- гликолипиды;
- холестерол.
К первой группе фосфолипидов относятся такие молекулы, как глицерофосфолипиды и сфингомиелины. Эти молекулы составляют основу бислоя мембраны. Гидрофобные концы соединений направлены внутрь слоя, гидрофильные — наружу. Примеры соединений:
- фосфатидилхолин;
- фосфатидилсерин;
- кардиолипин;
- фосфатидилинозитол;
- сфингомиелин;
- фосфатидилглицерин;
- фосфатидилэтаноламин.
Для изучения данных молекул применяется способ разрушения слоя мембраны в некоторых частях фосфолипазой — специальным ферментом, катализирующим процесс распада фосфолипидов.
Функции перечисленных соединений следующие:
- Обеспечивают общую структуру и строение бислоя плазмалеммы.
- Соприкасаются с белками на поверхности и внутри слоя.
- Определяют агрегатное состояние, которое будет иметь плазматическая мембрана клетки при различных температурных условиях.
- Участвуют в ограниченной проницаемости плазмолеммы для разных молекул.
- Формируют разные типы взаимодействий клеточных мембран друг с другом (десмосома, щелевидное пространство, плотный контакт).
Сфингофосфолипиды и гликолипиды мембраны
Сфингомиелины или сфингофосфолипиды по своей химической природе — производные аминоспирта сфингозина. Наравне с фосфолипидами принимают участие в образовании билипидного слоя мембраны.
К гликолипидам относится гликокаликс — вещество, во многом определяющее свойства плазматической мембраны. Это желеподобное соединение, состоящее в основном из олигосахаридов. Гликокаликс занимает 10% от общей массы плазмалеммы. С этим веществом напрямую связана плазматическая мембрана, строение и функции, которые она выполняет. Так, например, гликокаликс осуществляет:
- маркерную функцию мембраны;
- рецепторную;
- процессы пристеночного переваривания частиц внутри клетки.
Следует заметить, что наличие липида гликокаликса характерно только для животных клеток, но не для растительных, бактериальных и грибов.
Холестерол (стерин мембраны)
Является важной составной частью бислоя клетки у млекопитающих животных. В растительных не встречается, в бактериальных и грибах тоже. С химической точки зрения представляет собой спирт, циклический, одноатомный.
Равно как и остальные липиды, обладает свойствами амфифильности (наличие гидрофильного и гидрофобного конца молекулы). В мембране играет важную роль ограничителя и контролера текучести бислоя. Также участвует в выработке витамина D, является соучастником формирования половых гормонов.
В растительных же клетках присутствуют фитостеролы, которые не участвуют в образовании животных мембран. По некоторым данным известно, что эти вещества обеспечивают устойчивость растений к некоторым видам заболеваний.
Плазматическая мембрана образована холестеролом и другими липидами в общем взаимодействии, комплексе.
Углеводы мембраны
Данная группа веществ составляет примерно около 10% от общего состава соединений плазмалеммы. В простом виде моно-, ди-, полисахариды не встречаются, а только в форме гликопротеидов и гликолипидов.
Функции их заключаются в осуществлении контроля над внутри- и межклеточными взаимодействиями, поддержании определенной структуры и положения молекул белков в мембране, а также осуществлении рецепции.
Основные функции плазмалеммы
Очень велика роль, которую играет в клетке плазматическая мембрана. Функции ее многогранны и важны. Рассмотрим их подробнее.
- Отграничивает содержимое клетки от окружающей среды и защищает его от внешних воздействий. Благодаря наличию мембраны поддерживается на постоянном уровне химический состав цитоплазмы, ее содержимое.
- Плазмалемма содержит ряд белков, углеводов и липидов, которые придают и поддерживают определенную форму клетки.
- Мембрану имеет каждая клеточная органелла, которая называется мембранной везикулой (пузырьком).
- Компонентный состав плазмалеммы позволяет ей исполнять роль «стражника» клетки, осуществляя выборочный транспорт внутрь нее.
- Рецепторы, ферменты, биологически активные вещества функционируют в клетке и проникают в нее, сотрудничают с ее поверхностной оболочкой только благодаря белкам и липидам мембраны.
- Через плазмалемму осуществляется транспортировка не только соединений различной природы, но и ионов, важных для жизнедеятельности (натрий, калий, кальций и другие).
- Мембрана поддерживает осмотическое равновесие вне и внутри клетки.
- При помощи плазмалеммы осуществляется перенос ионов и соединений различной природы, электронов, гормонов из цитоплазмы в органеллы.
- Через нее же происходит поглощение солнечного света в виде квантов и пробуждение сигналов внутри клетки.
- Именно данной структурой осуществляется генерация импульсов действия и покоя.
- Механическая защита клетки и ее структур от небольших деформаций и физических воздействий.
- Адгезия клеток, то есть сцепление, и удержание их рядом друг с другом также осуществляется благодаря мембране.
Очень тесно взаимосвязана клеточная плазмалемма и цитоплазма. Плазматическая мембрана находится в тесном контакте со всеми веществами и молекулами, ионами, которые проникают внутрь клетки и свободно располагаются в вязкой внутренней среде. Данные соединения пытаются проникнуть внутрь всех клеточных структур, но барьером служит как раз мембрана, которая способна осуществлять разные типы транспорта через себя. Либо вообще не пропускать некоторые типы соединений.

Типы транспорта через клеточный барьер
Транспорт через плазматическую мембрану осуществляется несколькими способами, которые объединяет одна общая физическая особенность — закон диффузии веществ.
- Пассивный транспорт или диффузия и осмос. Подразумевает свободное перемещение ионов и растворителя через мембрану по градиенту из области с высокой концентрацией в область с низкой. Не требует расхода энергии, так как протекает сам по себе. Так происходит действие натрий-калиевого насоса, смена кислорода и углекислого газа при дыхании, выход глюкозы в кровь и так далее. Очень распространено такое явление, как облегченная диффузия. Данный процесс подразумевает наличие какого-либо вещества-помощника, которое цепляет нужное соединение и протаскивает за собой по белковому каналу или через липидный слой внутрь клетки.
- Активный транспорт подразумевает затраты энергии на процессы поглощения и выведения через мембрану. Есть два основных способа: экзоцитоз — выведение молекул и ионов наружу. Эндоцитоз — захватывание и проведение внутрь клетки твердых и жидких частиц. В свою очередь, второй способ активного транспорта включает в себя две разновидности процесса. Фагоцитоз, который заключается в заглатывании везикулой мембраны твердых молекул, веществ, соединений и ионов и проведение их внутрь клетки. При протекании данного процесса образуются крупные везикулы. Пиноцитоз, напротив, заключается в поглощении капелек жидкостей, растворителей и других веществ и проведении их внутрь клетки. Он подразумевает формирование пузырьков малых размеров.
Оба процесса — пиноцитоз и фагоцитоз — играют большую роль не только в осуществлении транспорта соединений и жидкостей, но и в защите клетки от обломков отмерших клеток, микроорганизмов и вредных соединений. Можно сказать, что эти способы активного транспорта также являются и вариантами иммунологической защиты клетки и ее структур от разных опасностей.
www.syl.ru
Биологические мембраны — «Энциклопедия»
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ (латинский membrana — кожица, оболочка, перепонка), структуры, ограничивающие содержимое клеток (клеточная, или плазматическая, мембрана, плазмалемма) и внутриклеточных органелл. У прокариот имеется только клеточная мембрана, в большинстве случаев окружённая клеточной стенкой. У эукариот мембраной окружена не только клетка, но и ядро, а также митохондрии, лизосомы, пероксисомы, секреторные гранулы, эндосомы, у растений ещё — хлоропласты и вакуоли; мембраны образуют также разветвлённую сеть эндоплазматического ретикулума и комплекса Гольджи. Митохондрии, хлоропласты и ядра окружены двумя мембранами, а внутри хлоропластов имеется ещё один тип мембран, формирующих тилакоиды. У животных к клеточной мембране снаружи примыкает гликопротеиновый комплекс — гликокаликс, у растений — клеточная стенка. Толщина мембран варьирует от 6 до 10 нм.
 Структура биологических мембран. Основу биологических мембран составляет протяжённый двойной слой (бислой) глицерофосфо-, сфинго- и гликолипидов со встроенными в него молекулами различных белков. Гидрофобные (неполярные) группы молекул липидов (остатки жирных кислот) погружены в толщу мембраны, а гидрофильные (полярные) головки ориентированы наружу, в окружающую водную среду (смотри Липиды). Плотность упаковки биологических мембран обеспечивается электростатическими взаимодействиями полярных головок и гидрофобными контактами между цепями жирных кислот. Входящие в состав биологических мембран белки взаимодействуют с липидным бислоем с помощью гидрофобных взаимодействий и вандерваальсовых связей. Соотношение липидов и белков, их состав в различных биологических мембранах могут существенно различаться. Так, в мембранах миелиновой оболочки содержание липидов (по массе) в четыре раза больше, чем белков, а во внутренних мембранах митохондрий более чем в два раза преобладают белки. Липиды биологических мембран представлены главным образом фосфатидилхолином, фосфатидилэтаноламином, сфингомиелином, фосфатидилсерином, фосфатидилинозитом и кардиолипином, которые обнаруживаются примерно в одном и том же соотношении в мембранах разных по уровню организации организмов. В то же время набор жирных кислот, входящих в состав липидов, подвержен изменениям. Например, понижение температуры, давления и солёности среды обитания организмов сопровождаются увеличением количества ненасыщенных связей и/или короткоцепочечных жирных кислот в фосфолипидах и происходящим вследствие этого уменьшением плотности упаковки бислоя. Липидам свойственна определённая подвижность внутри бислоя. Они способны к быстрому вращению вокруг оси (вращательная диффузия), к свободному перемещению в пределах одного слоя мембраны (латеральная диффузия), а также к переходу с одной стороны бислоя на другую (такое передвижение обеспечивается специальными механизмами). Для клеточных мембран животных клеток характерно высокое содержание холестерина (в среднем около 21%), который участвует в регуляции текучести мембраны, препятствуя плотной упаковке фосфолипидов. В растительной клетке роль холестерина играет его аналог — десмостерин. В мембранах бактерий и внутриклеточных органелл стерины отсутствуют. До 10% сухого вещества мембран приходится на долю углеводов, которые экспонированы на внешней стороне клеточной мембраны и являются составной частью мембранных гликолипидов и гликопротеинов.
Структура биологических мембран. Основу биологических мембран составляет протяжённый двойной слой (бислой) глицерофосфо-, сфинго- и гликолипидов со встроенными в него молекулами различных белков. Гидрофобные (неполярные) группы молекул липидов (остатки жирных кислот) погружены в толщу мембраны, а гидрофильные (полярные) головки ориентированы наружу, в окружающую водную среду (смотри Липиды). Плотность упаковки биологических мембран обеспечивается электростатическими взаимодействиями полярных головок и гидрофобными контактами между цепями жирных кислот. Входящие в состав биологических мембран белки взаимодействуют с липидным бислоем с помощью гидрофобных взаимодействий и вандерваальсовых связей. Соотношение липидов и белков, их состав в различных биологических мембранах могут существенно различаться. Так, в мембранах миелиновой оболочки содержание липидов (по массе) в четыре раза больше, чем белков, а во внутренних мембранах митохондрий более чем в два раза преобладают белки. Липиды биологических мембран представлены главным образом фосфатидилхолином, фосфатидилэтаноламином, сфингомиелином, фосфатидилсерином, фосфатидилинозитом и кардиолипином, которые обнаруживаются примерно в одном и том же соотношении в мембранах разных по уровню организации организмов. В то же время набор жирных кислот, входящих в состав липидов, подвержен изменениям. Например, понижение температуры, давления и солёности среды обитания организмов сопровождаются увеличением количества ненасыщенных связей и/или короткоцепочечных жирных кислот в фосфолипидах и происходящим вследствие этого уменьшением плотности упаковки бислоя. Липидам свойственна определённая подвижность внутри бислоя. Они способны к быстрому вращению вокруг оси (вращательная диффузия), к свободному перемещению в пределах одного слоя мембраны (латеральная диффузия), а также к переходу с одной стороны бислоя на другую (такое передвижение обеспечивается специальными механизмами). Для клеточных мембран животных клеток характерно высокое содержание холестерина (в среднем около 21%), который участвует в регуляции текучести мембраны, препятствуя плотной упаковке фосфолипидов. В растительной клетке роль холестерина играет его аналог — десмостерин. В мембранах бактерий и внутриклеточных органелл стерины отсутствуют. До 10% сухого вещества мембран приходится на долю углеводов, которые экспонированы на внешней стороне клеточной мембраны и являются составной частью мембранных гликолипидов и гликопротеинов.
Реклама
Содержание белка в различных мембранах колеблется от 20 до 75% (в пересчёте на сухую массу). Мембранные белки могут быть встроены в бислой (интегральные белки). При этом они погружены в мембрану и пронизывают её (иногда несколько раз) таким образом, что достаточно протяжённые участки белка, образованные гидрофобными аминокислотами, оказываются в её толще, а гидрофильные — на поверхности, по обе стороны биологических мембран. Выступающие над внешней стороной мембраны участки белковых молекул обычно несут несколько ковалентно связанных, часто разветвлённых цепей олигосахаридов, образованных остатками маннозы, фукозы, глюкозы, N-ацетилглюкозамина и др. Эти компоненты играют роль маркеров при распознавании клеточной поверхности. Молекулы периферических белков расположены главным образом на внутренней поверхности мембраны, не проникая внутрь бислоя, и удерживаются на ней с помощью электростатических взаимодействий и водородных связей; они связываются с мембраной обратимо и могут переходить в цитоплазму при модификации белков (например, путём их фосфорилирования) в ответ на изменения функционального состояния клетки. Многие белки организованы в виде сложных комплексов (например, белки дыхательной цепи митохондрий). В клетках происходит постоянное обновление компонентов биологических мембран путём введения новых молекул липидов и белков, однако структурная организация биологических мембран в течение всей жизни клетки остаётся неизменной.
Функции мембран. Основные функции мембран связаны с белками. Многие мембранные белки — ферменты, обеспечивающие протекание окислительно-восстановительных, гидролитических и биосинтетических реакций как на поверхности мембраны, так и внутри неё. Важнейшая функция белков мембран — транспортная. Жирорастворимые соединения (например, стероидные гормоны) легко проникают в липидный бислой, но для большинства других соединений (в том числе аминокислот, сахаров) и неорганических ионов он непроницаем. Специально предназначенные для этих целей мембранные белки обеспечивают как активный (требующий затрат энергии), так и пассивный (за счёт градиента концентраций) транспорт веществ и ионов (смотри Ионные каналы). Специфические белки — ионные насосы (Na/К-насос и Са-насос) ответственны за асимметричное распределение ионов Na+, К+ и Са2+ по обе стороны клеточной мембраны (например, в цитоплазме животной клетки — низкую для Na+ и Са2+ и высокую для К+). Такая асимметрия обеспечивает многие проявления жизнедеятельности (электровозбудимость, осмотическую устойчивость и др.). Мембранные белки аквапорины образуют в мембране специальные каналы, регулирующие проникновение в клетку молекул воды. Взаимодействие клетки с внешней средой, регуляция внутриклеточных процессов осуществляются посредством рецепторных белков (рецепторов), ответственных за фото-, термо-, механо- и хеморецепцию.
 Барьерная функция биологических мембран обеспечивает сохранение определённого состава клетки и концентрации составляющих её веществ, а также защиту от воздействия различных чужеродных факторов и токсинов. Благодаря биологическим мембранам внутри клеток возможно одновременное протекание множества несовместимых друг с другом реакций. Например, необходимые клетке белки синтезируются на рибосомах, прикреплённых к эндоплазматическому ретикулуму, а их распад происходит в лизосомах. В биологических мембранах протекают процессы энергообмена клеток. Внутренние мембраны митохондрий и мембраны тилакоидов — важнейшие преобразователи энергии, играющие ключевую роль в запасании энергии, образующейся в ходе дыхания и фотосинтеза, в энергию пирофосфатной связи аденозинтрифосфата. Биологические мембраны нейронов могут генерировать и осуществлять передачу электрического сигнала, участвуя тем самым в процессах возбуждения и проведения нервного импульса.
Барьерная функция биологических мембран обеспечивает сохранение определённого состава клетки и концентрации составляющих её веществ, а также защиту от воздействия различных чужеродных факторов и токсинов. Благодаря биологическим мембранам внутри клеток возможно одновременное протекание множества несовместимых друг с другом реакций. Например, необходимые клетке белки синтезируются на рибосомах, прикреплённых к эндоплазматическому ретикулуму, а их распад происходит в лизосомах. В биологических мембранах протекают процессы энергообмена клеток. Внутренние мембраны митохондрий и мембраны тилакоидов — важнейшие преобразователи энергии, играющие ключевую роль в запасании энергии, образующейся в ходе дыхания и фотосинтеза, в энергию пирофосфатной связи аденозинтрифосфата. Биологические мембраны нейронов могут генерировать и осуществлять передачу электрического сигнала, участвуя тем самым в процессах возбуждения и проведения нервного импульса.
Белковые и липидные компоненты выполняют ряд других функций. Фрагменты фосфолипидов могут выступать в качестве предшественников сигнальных молекул (мессенджеров). Например, при активации мембранной фосфолипазы А из бислоя высвобождается арахидоновая кислота, дальнейшие превращения которой приводят к образованию биологических регуляторов — тромбоксанов, лейкотриенов и простагландинов. Фосфатидилсерин, локализованный на внутренней стороне мембраны, при инициации апоптоза мигрирует на её внешнюю сторону. Его появление служит сигналом для фагоцитов, которые имеют рецепторы на этот фосфолипид; они «узнают» дефектные клетки и уничтожают их. Гликолипиды наряду с гликопротеинами играют важную роль в явлениях межклеточной адгезии, участвуют в иммунных реакциях.
Методы изучения биологических мембран. Для изучения структуры и функции мембран используются электрофизиологические и иммуноцитохимические методы, жидкостная хроматография (для идентификации и анализа липидных компонентов), проточная цитометрия, позволяющая проследить ответ клетки на взаимодействие специфических лигандов с клеточной мембраной, разнообразные физические методы, характеризующие структуру мембран, упаковку и подвижность липидов в бислое (в том числе электронная микроскопия, малоугловое рассеивание нейтронов, флуоресцентная спектроскопия, круговой дихроизм), и другие методы.
Разнообразие типов биологических мембран, их полифункциональность и высокая чувствительность к внешним воздействиям являются причиной того, что они вовлекаются в различные патологические процессы. Повреждения клеточных мембран, приводящие к образованию свободных радикалов и гибели нервных клеток, лежат в основе нейродегенеративных заболеваний (болезнь Альцгеймера, паркинсонизм, боковой амиотрофический склероз), могут служить причиной инсульта и инфаркта миокарда.
Лит.: Крепс Е.М. Липиды клеточных мембран. Л., 1981; Введение в биомембранологию. М., 1990; Биологические мембраны. Методы. М., 1990; Геннис Р. Биологические мембраны. М., 1999; Пальцев М. А., Иванов А. А., Северин С. Е. Межклеточные взаимодействия. 2-е изд. М., 2003; Ченцов Ю. С. Введение в клеточную биологию. 4-е изд. М., 2004.
А. А. Болдырев.
knowledge.su